

17beta-Estradiol-induced enhancement of estrogen receptor biosynthesis via MAPK pathway in mouse skeletal muscle myoblasts
- PMID: 19340542
- PMCID: PMC10717592
- DOI: 10.1007/s12576-009-0023-0
17beta-Estradiol-induced enhancement of estrogen receptor biosynthesis via MAPK pathway in mouse skeletal muscle myoblasts
Abstract
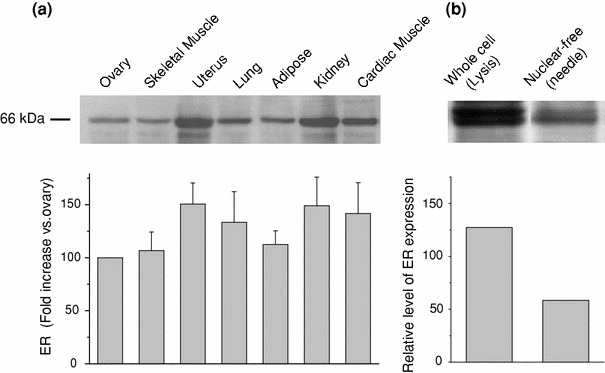
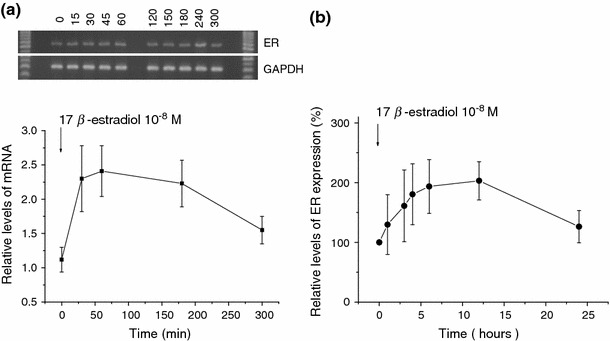
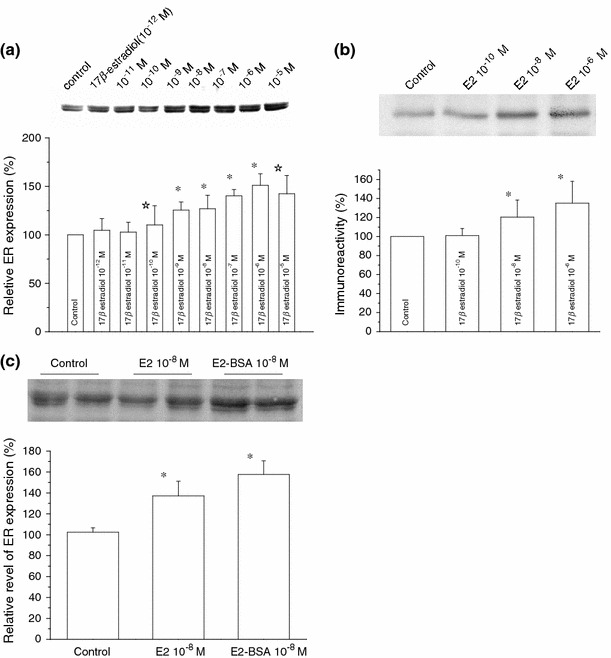
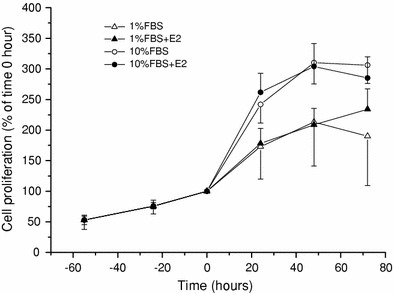
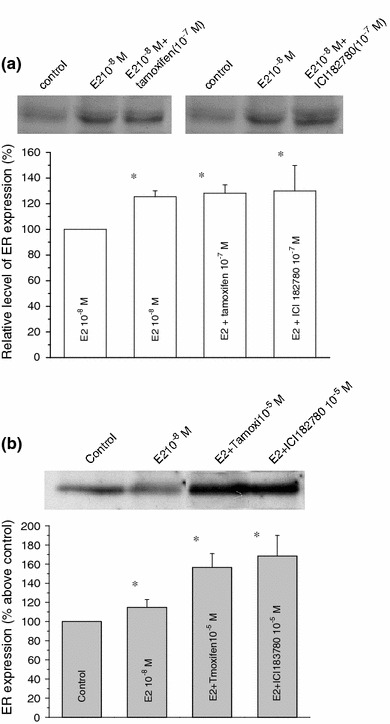
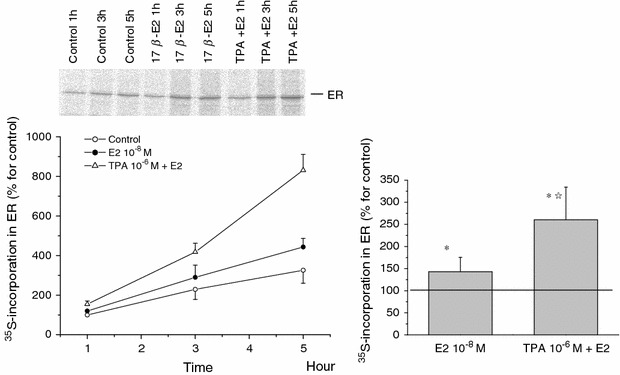
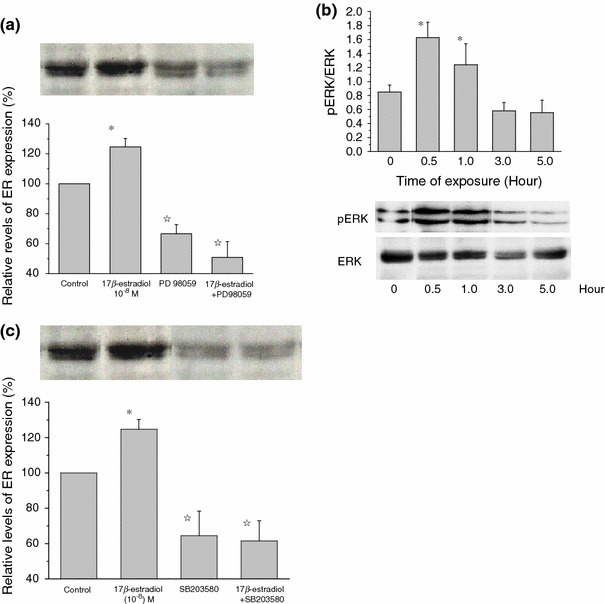
The skeletal muscle is one of the important target tissues for the actions of estrogen via both nuclear and extranuclear (non-genomic) pathways. However, there is a paucity of information about the receptor (ER) involved. The aim of this study was thus to explore the ER expression in skeletal muscle, and the influence of estrogen on it, by using C2C12 myoblasts derived from mouse skeletal muscle. Significant expression of a approximately 66-kD protein immunoreactive to ER type alpha (ERalpha) monoclonal antibody, which was comparable to that in ovary, was detected in the whole-cell (total) and nucleus-free (nonnuclear) fractions of C2C12 myoblasts. The expression level of these ER proteins increased in several hours with treatment with 17beta-estradiol (E2), which was preceded by the elevation of the ER mRNA level. This increase appeared to reflect the acceleration of de novo synthesis of ER protein, as proved by the (35)S-methionine immunoprecipitation method. A similar extent of fast increase in ER expression was also induced by a membrane-impermeable, BSA-conjugated estradiol (E2-BSA). Unexpectedly, the E2-induced increases in total and nonnuclear ER were further enhanced by the classic ER antagonists tamoxifen and ICI182,780 in a wide concentration range, implying some structural difference of the involved ER from the classical one. Treatment with the ERK1/2 inhibitor, PD98059 (10 microM), or the p38 MAPK-specific inhibitor, SB203580 (10 microM), greatly inhibited the E2-induced ER increase, while the protein kinase C (PKC) activator TPA (1 microM) enhanced it. These results collectively suggest that C2C12 skeletal myoblasts express a high level of ER, a considerable part of which is extranuclear. Further, the expression of ER in these cells may be significantly upregulated by estrogen itself via increased biosynthesis linked to membrane-bound ER and downstream MAPK-mediated signaling pathways.
Figures
References
-
- Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson G, Enmark E, Pettersson K, Warner M, Gustafsson JA. Mechanisms of estrogen action. Physiol Rev. 2001;81:1535–1565. - PubMed
-
- Nadal A, Diaz M, Valverde M. The estrogen trinity: membrane, cytosolic, and nuclear effects. News Physiol Sci. 2001;16:251–255. - PubMed
-
- Acconcia F, Ascenzi P, Fabozzi G, Visca P, Marino M. S-palmitoylation modulates human estrogen receptor-α functions. BBRC. 2004;316:878–883. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
