

Small molecules efficiently direct endodermal differentiation of mouse and human embryonic stem cells
- PMID: 19341624
- PMCID: PMC4564293
- DOI: 10.1016/j.stem.2009.01.014
Small molecules efficiently direct endodermal differentiation of mouse and human embryonic stem cells
Abstract
An essential step for therapeutic and research applications of stem cells is the ability to differentiate them into specific cell types. Endodermal cell derivatives, including lung, liver, and pancreas, are of interest for regenerative medicine, but efforts to produce these cells have been met with only modest success. In a screen of 4000 compounds, two cell-permeable small molecules were indentified that direct differentiation of ESCs into the endodermal lineage. These compounds induce nearly 80% of ESCs to form definitive endoderm, a higher efficiency than that achieved by Activin A or Nodal, commonly used protein inducers of endoderm. The chemically induced endoderm expresses multiple endodermal markers, can participate in normal development when injected into developing embryos, and can form pancreatic progenitors. The application of small molecules to differentiate mouse and human ESCs into endoderm represents a step toward achieving a reproducible and efficient production of desired ESC derivatives.
Figures

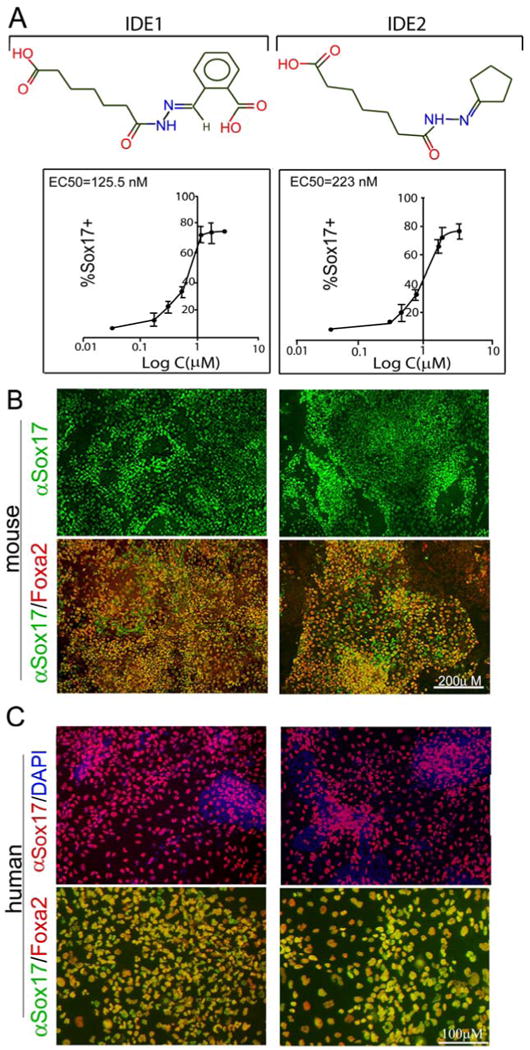




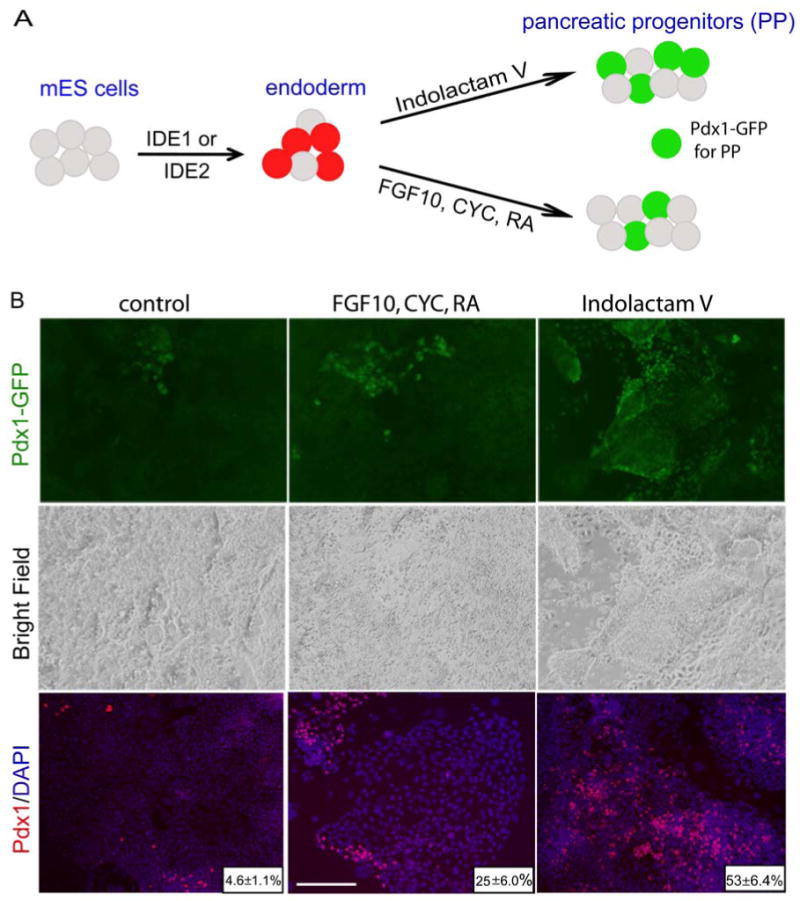
Comment in
-
Using small molecules to great effect in stem cell differentiation.Cell Stem Cell. 2009 May 8;4(5):373-4. doi: 10.1016/j.stem.2009.04.012. Cell Stem Cell. 2009. PMID: 19427285
References
-
- Ang SL, Wierda A, Wong D, Stevens KA, Cascio S, Rossant J, Zaret KS. The formation and maintenance of the definitive endoderm lineage in the mouse: involvement of HNF3/forkhead proteins. Development. 1993;119:1301–1315. - PubMed
-
- Bhushan A, Itoh N, Kato S, Thiery JP, Czernichow P, Bellusci S, Scharfmann R. Fgf10 is essential for maintaining the proliferative capacity of epithelial progenitor cells during early pancreatic organogenesis. Development. 2001;128:5109–5117. - PubMed
-
- Burridge PW, Anderson D, Priddle H, Barbadillo Munoz MD, Chamberlain S, Allegrucci C, Young LE, Denning C. Improved human embryonic stem cell embryoid body homogeneity and cardiomyocyte differentiation from a novel V-96 plate aggregation system highlights interline variability. Stem Cells. 2007;25:929–938. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
