

Inhibition of autoregulated TGFbeta signaling simultaneously enhances proliferation and differentiation of kidney epithelium and promotes repair following renal ischemia
- PMID: 19342372
- PMCID: PMC2671361
- DOI: 10.2353/ajpath.2009.080295
Inhibition of autoregulated TGFbeta signaling simultaneously enhances proliferation and differentiation of kidney epithelium and promotes repair following renal ischemia
Abstract
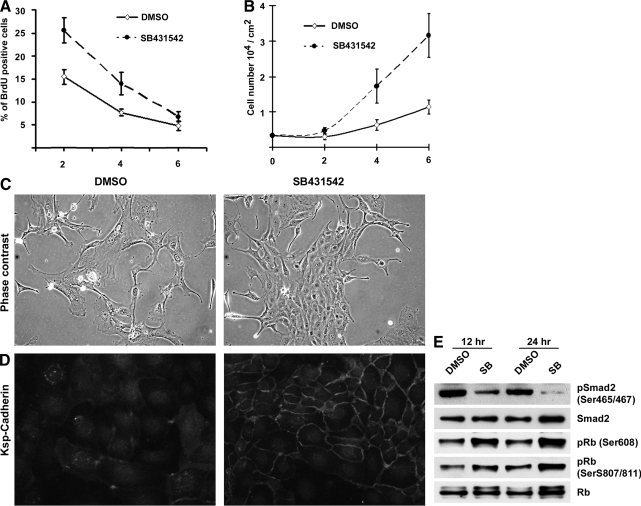
We studied autocrine transforming growth factor (TGF)beta signaling in kidney epithelium. Cultured proximal tubule cells showed regulated signaling that was high during log-phase growth, low during contact-inhibited differentiation, and rapidly increased during regeneration of wounded epithelium. Autoregulation of signaling correlated with TGFbeta receptor and Smad7 levels, but not with active TGFbeta, which was barely measurable in the growth medium. Confluent differentiated cells with low receptor and high Smad7 levels exhibited blunted responses to saturating concentrations of exogenously provided active TGFbeta, suggesting that TGFbeta signaling homeostasis was achieved by cell density-dependent modulation of signaling intermediates. Antagonism of Alk5 kinase, the TGFbeta type I receptor, dramatically accelerated the induction of differentiation in sparse, proliferating cultures and permitted better retention of differentiated features in regenerating cells of wounded, confluent cultures. Alk5 antagonism accelerated the differentiation of cells in proximal tubule primary cultures while simultaneously increasing their proliferation. Consequently, Alk5-inhibited primary cultures formed confluent, differentiated monolayers faster than untreated cultures. Furthermore, treatment with an Alk5 antagonist promoted kidney repair reflected by increased tubule differentiation and decreased tubulo-interstitial pathology during the recovery phase following ischemic injury in vivo. Our results show that autocrine TGFbeta signaling in proliferating proximal tubule cells exceeds the levels that are necessary for physiological regeneration. To that end, TGFbeta signaling is redundant and maladaptive during tubule repair by epithelial regeneration.
Figures










Similar articles
-
PTEN loss defines a TGF-β-induced tubule phenotype of failed differentiation and JNK signaling during renal fibrosis.Am J Physiol Renal Physiol. 2012 May 1;302(9):F1210-23. doi: 10.1152/ajprenal.00660.2011. Epub 2012 Feb 1. Am J Physiol Renal Physiol. 2012. PMID: 22301622 Free PMC article.
-
Activin Receptor-Like Kinase Receptors ALK5 and ALK1 Are Both Required for TGFβ-Induced Chondrogenic Differentiation of Human Bone Marrow-Derived Mesenchymal Stem Cells.PLoS One. 2015 Dec 31;10(12):e0146124. doi: 10.1371/journal.pone.0146124. eCollection 2015. PLoS One. 2015. PMID: 26720610 Free PMC article.
-
Activin receptor-like kinase (ALK)1 is an antagonistic mediator of lateral TGFbeta/ALK5 signaling.Mol Cell. 2003 Oct;12(4):817-28. doi: 10.1016/s1097-2765(03)00386-1. Mol Cell. 2003. PMID: 14580334
-
Tgf-beta type I receptor (Alk5) kinase inhibitors in oncology.Curr Pharm Biotechnol. 2011 Dec;12(12):2190-202. doi: 10.2174/138920111798808257. Curr Pharm Biotechnol. 2011. PMID: 21619541 Review.
-
TGFβ signaling in head and neck squamous cell carcinoma.Oncogene. 2010 Oct 7;29(40):5437-46. doi: 10.1038/onc.2010.306. Epub 2010 Aug 2. Oncogene. 2010. PMID: 20676130 Free PMC article. Review.
Cited by
-
Deleting the TGF-β receptor attenuates acute proximal tubule injury.J Am Soc Nephrol. 2012 Dec;23(12):2001-11. doi: 10.1681/ASN.2012020139. Epub 2012 Nov 15. J Am Soc Nephrol. 2012. PMID: 23160515 Free PMC article.
-
Persistent activation of autophagy in kidney tubular cells promotes renal interstitial fibrosis during unilateral ureteral obstruction.Autophagy. 2016 Jun 2;12(6):976-98. doi: 10.1080/15548627.2016.1166317. Epub 2016 Apr 28. Autophagy. 2016. PMID: 27123926 Free PMC article.
-
Conditional knockout of TGF-βRII /Smad2 signals protects against acute renal injury by alleviating cell necroptosis, apoptosis and inflammation.Theranostics. 2019 Oct 21;9(26):8277-8293. doi: 10.7150/thno.35686. eCollection 2019. Theranostics. 2019. PMID: 31754396 Free PMC article.
-
Autocrine transforming growth factor-{beta}1 activation mediated by integrin {alpha}V{beta}3 regulates transcriptional expression of laminin-332 in Madin-Darby canine kidney epithelial cells.Mol Biol Cell. 2010 Nov 1;21(21):3654-68. doi: 10.1091/mbc.E10-06-0523. Epub 2010 Sep 15. Mol Biol Cell. 2010. PMID: 20844080 Free PMC article.
-
Inhibition of Transforming Growth Factor-β Improves Primary Renal Tubule Cell Differentiation in Long-Term Culture.Tissue Eng Part A. 2023 Feb;29(3-4):102-111. doi: 10.1089/ten.TEA.2022.0147. Epub 2022 Nov 18. Tissue Eng Part A. 2023. PMID: 36274231 Free PMC article.
References
-
- Nony PA, Schnellmann RG. Mechanisms of renal cell repair and regeneration after acute renal failure. J Pharmacol Exp Ther. 2003;304:905–912. - PubMed
-
- Bonventre JV. Dedifferentiation and proliferation of surviving epithelial cells in acute renal failure, J Am Soc Nephrol. 2003;14 Suppl 1:S55–S61. - PubMed
-
- Devarajan P. Update on mechanisms of ischemic acute kidney injury. J Am Soc Nephrol. 2006;17:1503–1520. - PubMed
-
- Siegel PM, Massague J. Cytostatic and apoptotic actions of TGF-beta in homeostasis and cancer. Nat Rev Cancer. 2003;3:807–821. - PubMed
-
- Derynck R, Zhang YE. Smad-dependent and Smad-independent pathways in TGF-beta family signalling. Nature. 2003;425:577–584. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
