

Supramolecular assemblies and localized regulation of voltage-gated ion channels
- PMID: 19342611
- PMCID: PMC2733249
- DOI: 10.1152/physrev.00029.2007
Supramolecular assemblies and localized regulation of voltage-gated ion channels
Abstract
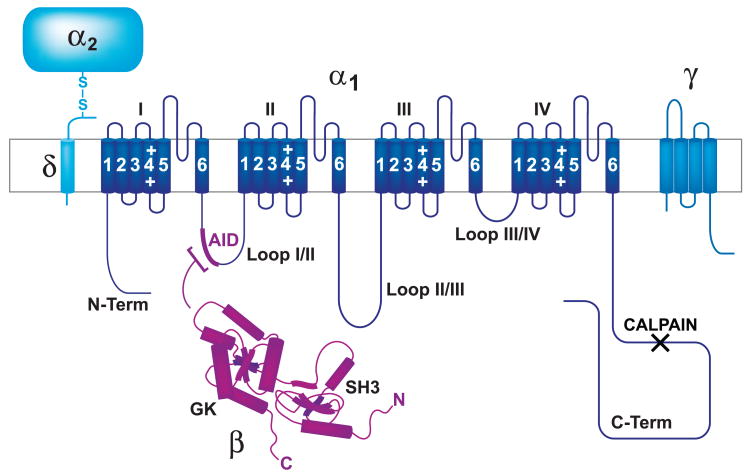
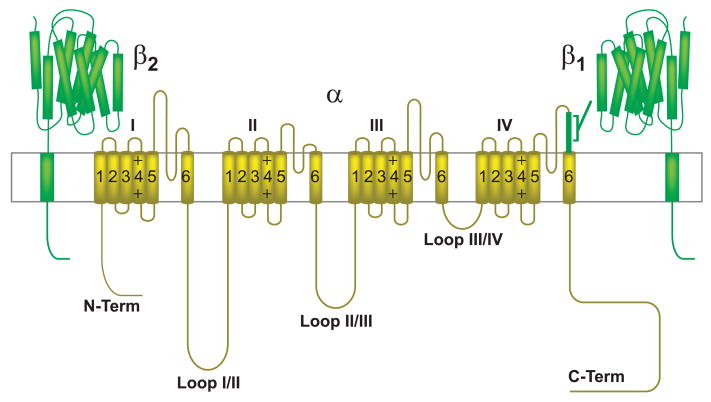
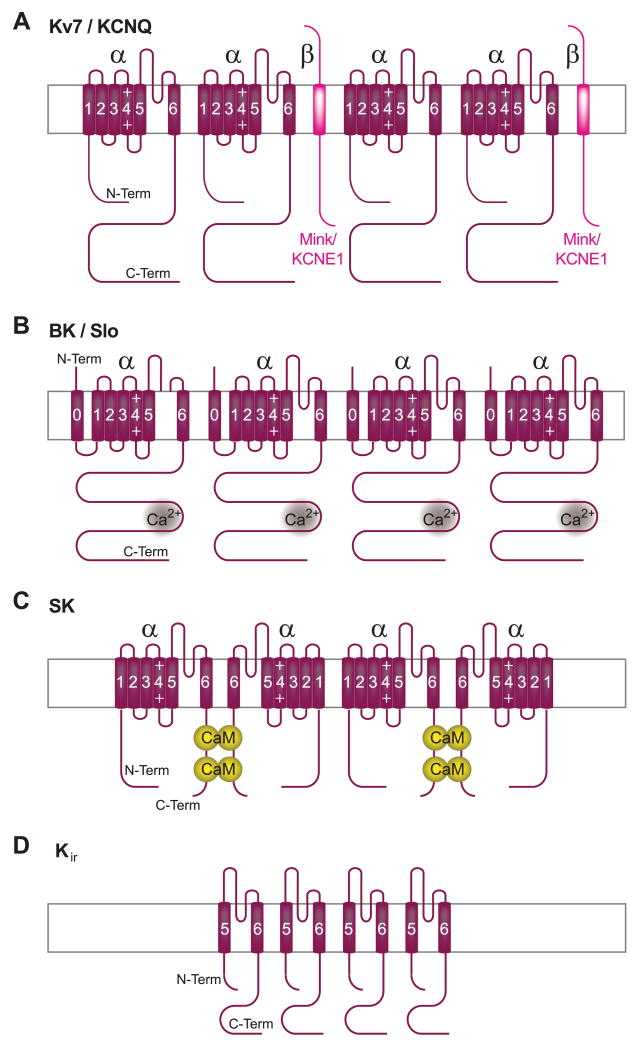
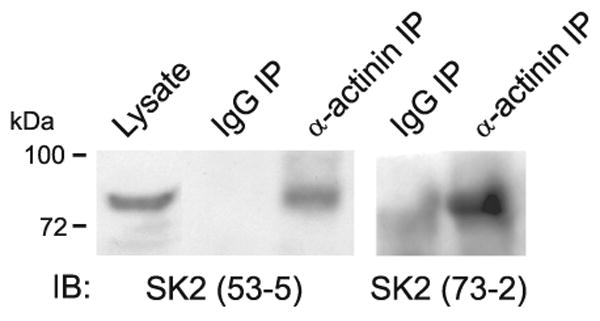
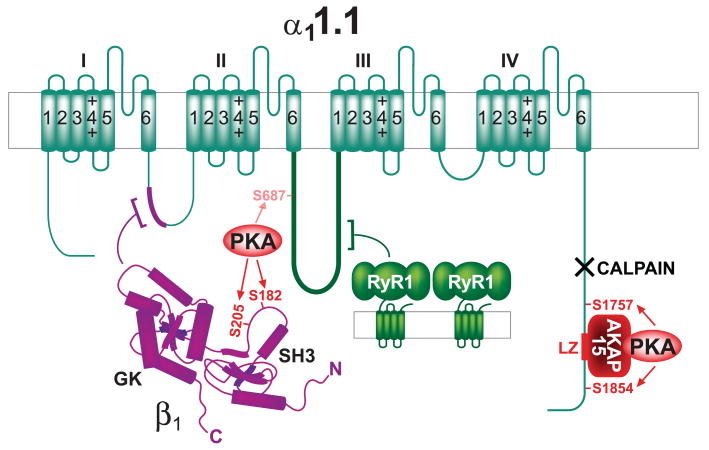
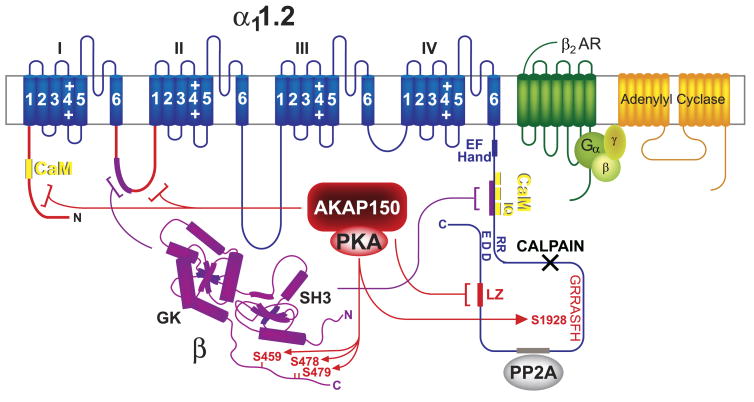
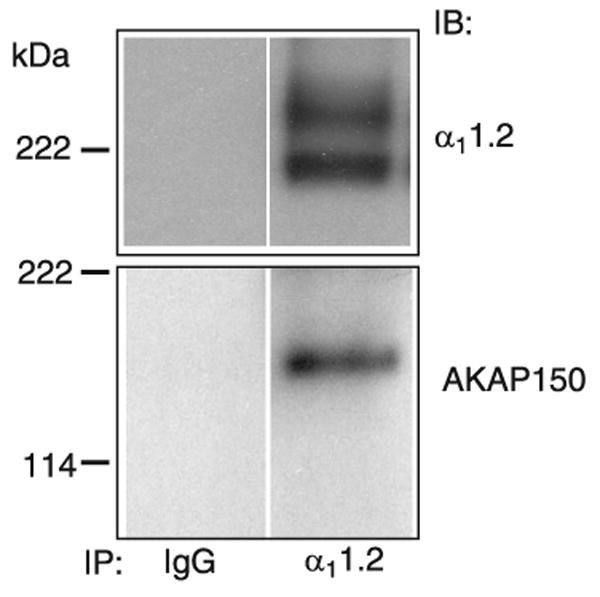
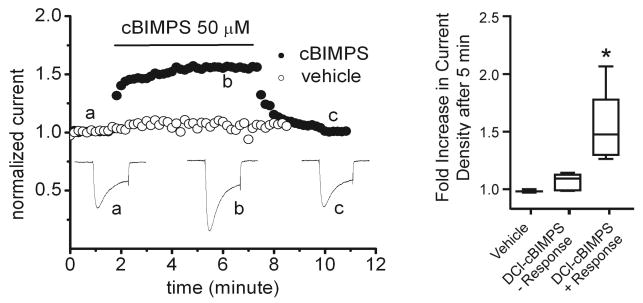
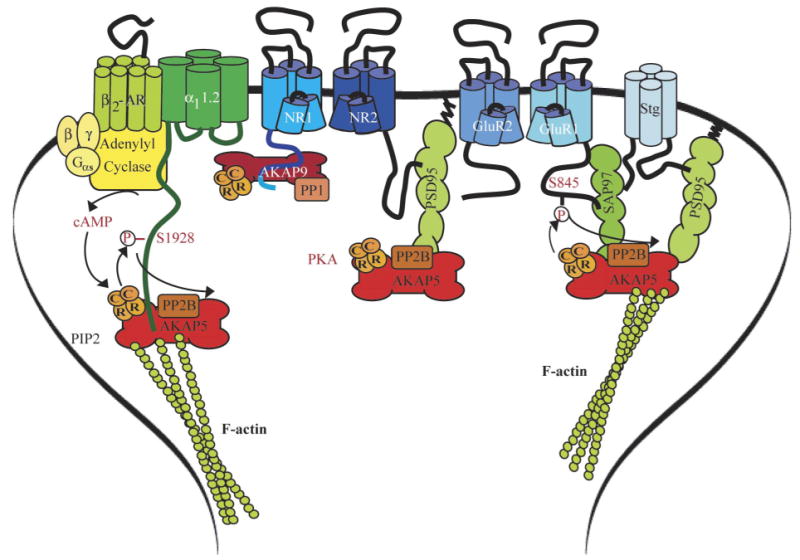
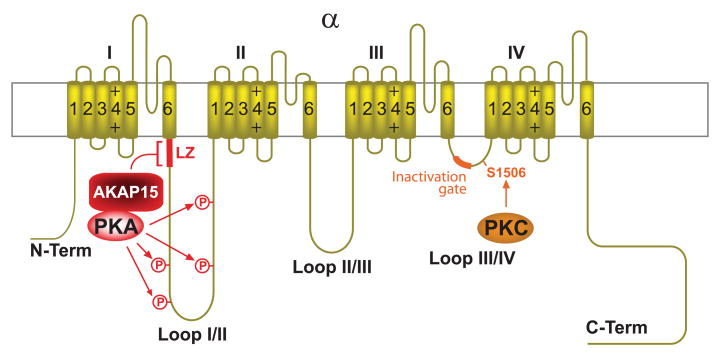
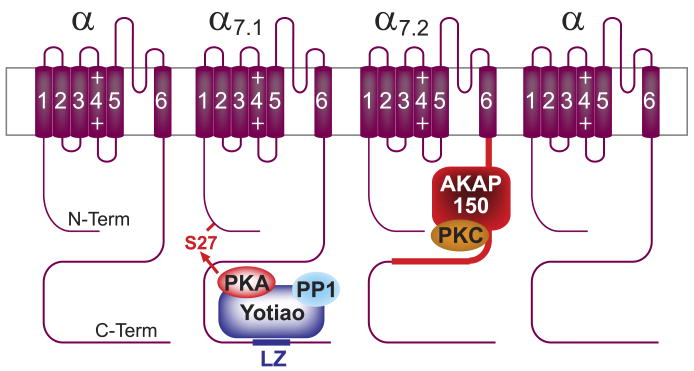
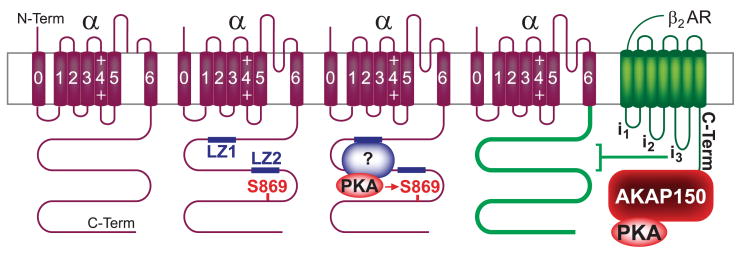
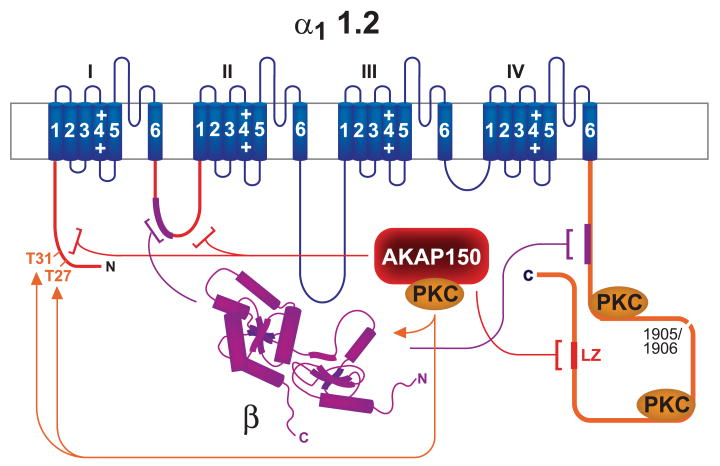
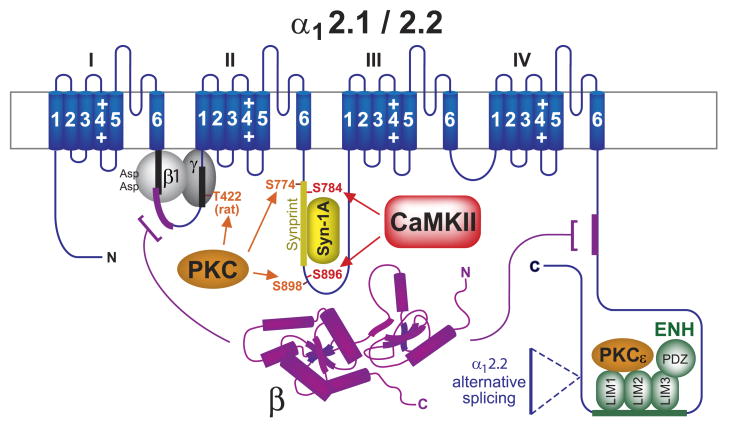
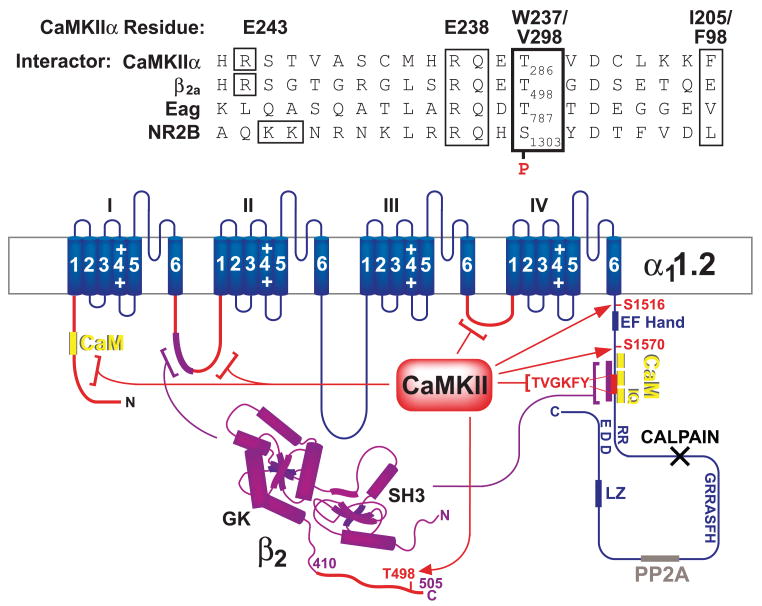
This review addresses the localized regulation of voltage-gated ion channels by phosphorylation. Comprehensive data on channel regulation by associated protein kinases, phosphatases, and related regulatory proteins are mainly available for voltage-gated Ca2+ channels, which form the main focus of this review. Other voltage-gated ion channels and especially Kv7.1-3 (KCNQ1-3), the large- and small-conductance Ca2+-activated K+ channels BK and SK2, and the inward-rectifying K+ channels Kir3 have also been studied to quite some extent and will be included. Regulation of the L-type Ca2+ channel Cav1.2 by PKA has been studied most thoroughly as it underlies the cardiac fight-or-flight response. A prototypical Cav1.2 signaling complex containing the beta2 adrenergic receptor, the heterotrimeric G protein Gs, adenylyl cyclase, and PKA has been identified that supports highly localized via cAMP. The type 2 ryanodine receptor as well as AMPA- and NMDA-type glutamate receptors are in close proximity to Cav1.2 in cardiomyocytes and neurons, respectively, yet independently anchor PKA, CaMKII, and the serine/threonine phosphatases PP1, PP2A, and PP2B, as is discussed in detail. Descriptions of the structural and functional aspects of the interactions of PKA, PKC, CaMKII, Src, and various phosphatases with Cav1.2 will include comparisons with analogous interactions with other channels such as the ryanodine receptor or ionotropic glutamate receptors. Regulation of Na+ and K+ channel phosphorylation complexes will be discussed in separate papers. This review is thus intended for readers interested in ion channel regulation or in localization of kinases, phosphatases, and their upstream regulators.
Figures
References
-
- Altier C, Dubel SJ, Barrere C, Jarvis SE, Stotz SC, Spaetgens RL, Scott JD, Cornet V, De Waard M, Zamponi GW, Nargeot J, Bourinet E. Trafficking of L-type calcium channels mediated by the postsynaptic scaffolding protein AKAP79. J Biol Chem. 2002;277:33598–33603. - PubMed
-
- Altier C, Khosravani H, Evans RM, Hameed S, Peloquin JB, Vartian BA, Chen L, Beedle AM, Ferguson SS, Mezghrani A, Dubel SJ, Bourinet E, McRory JE, Zamponi GW. ORL1 receptor-mediated internalization of N-type calcium channels. Nat Neurosci. 2006;9:31–40. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous