

Group V secretory phospholipase A2 modulates phagosome maturation and regulates the innate immune response against Candida albicans
- PMID: 19342668
- PMCID: PMC2746418
- DOI: 10.4049/jimmunol.0803776
Group V secretory phospholipase A2 modulates phagosome maturation and regulates the innate immune response against Candida albicans
Abstract
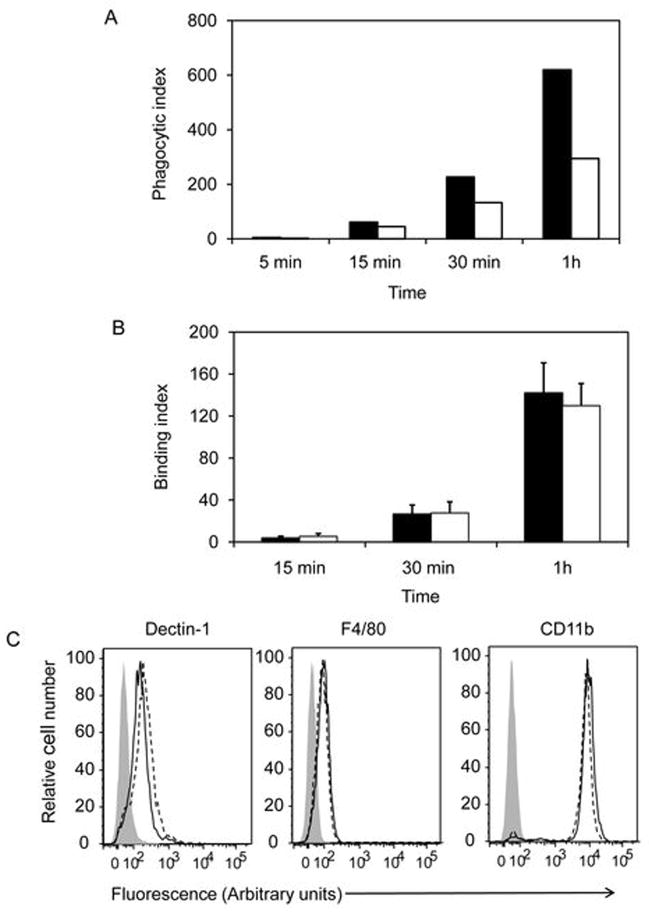
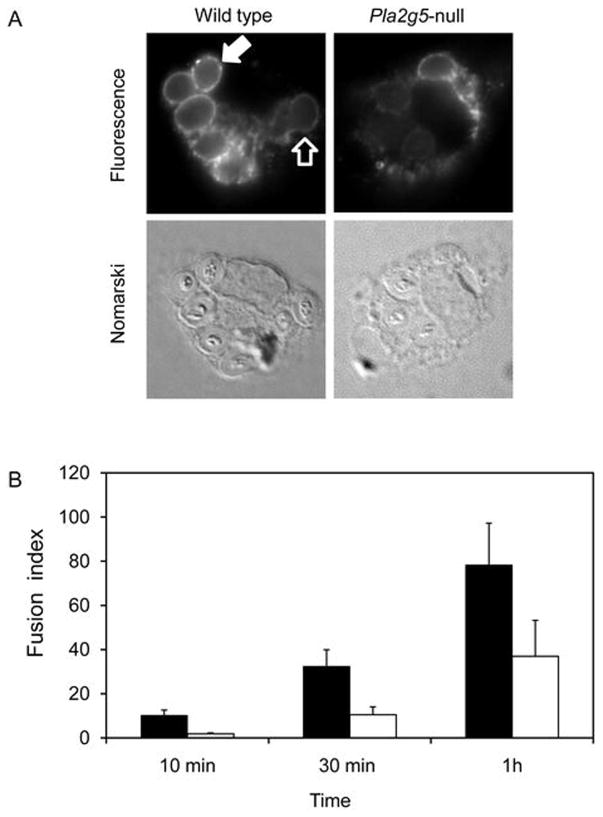
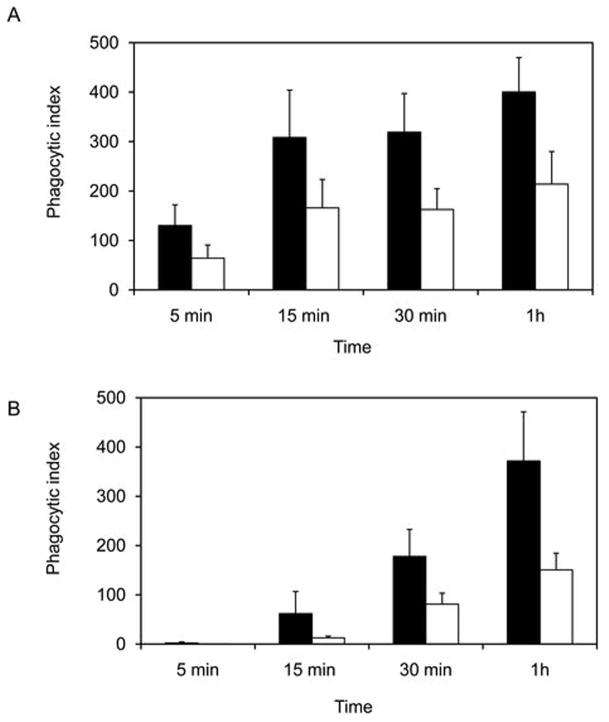
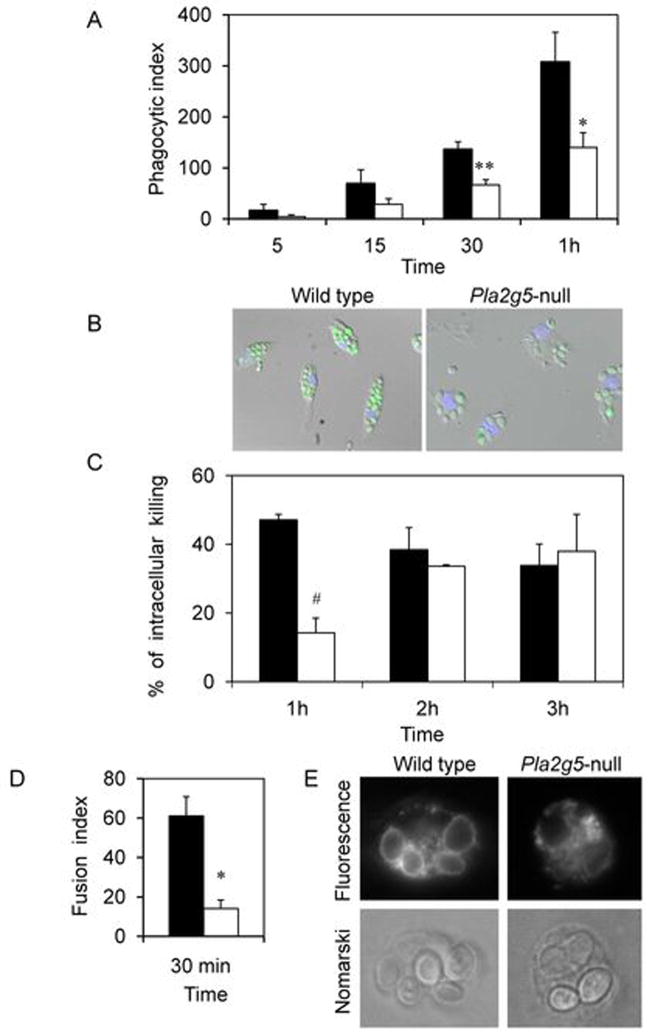
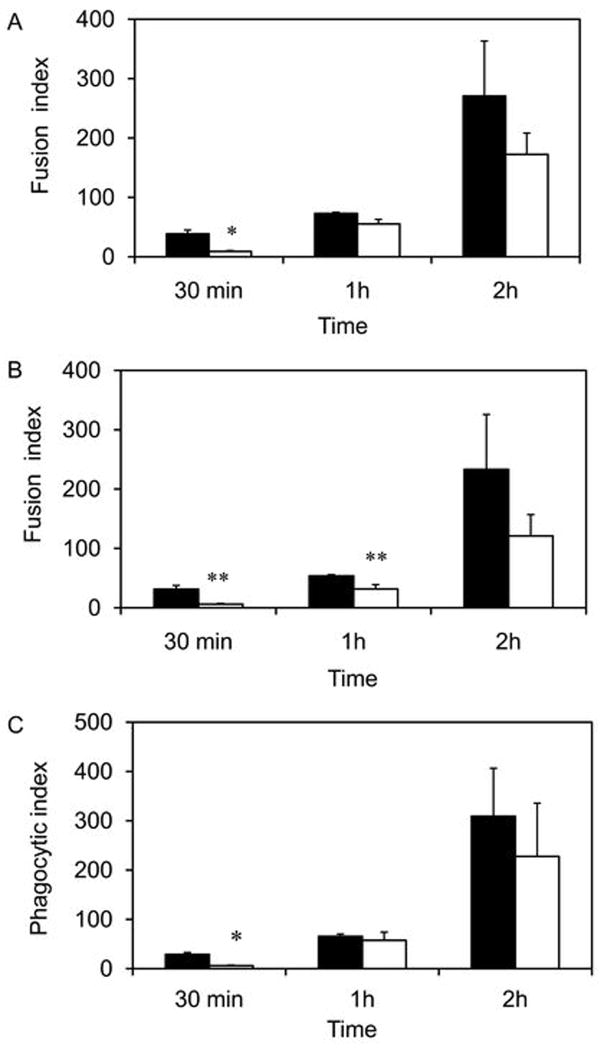
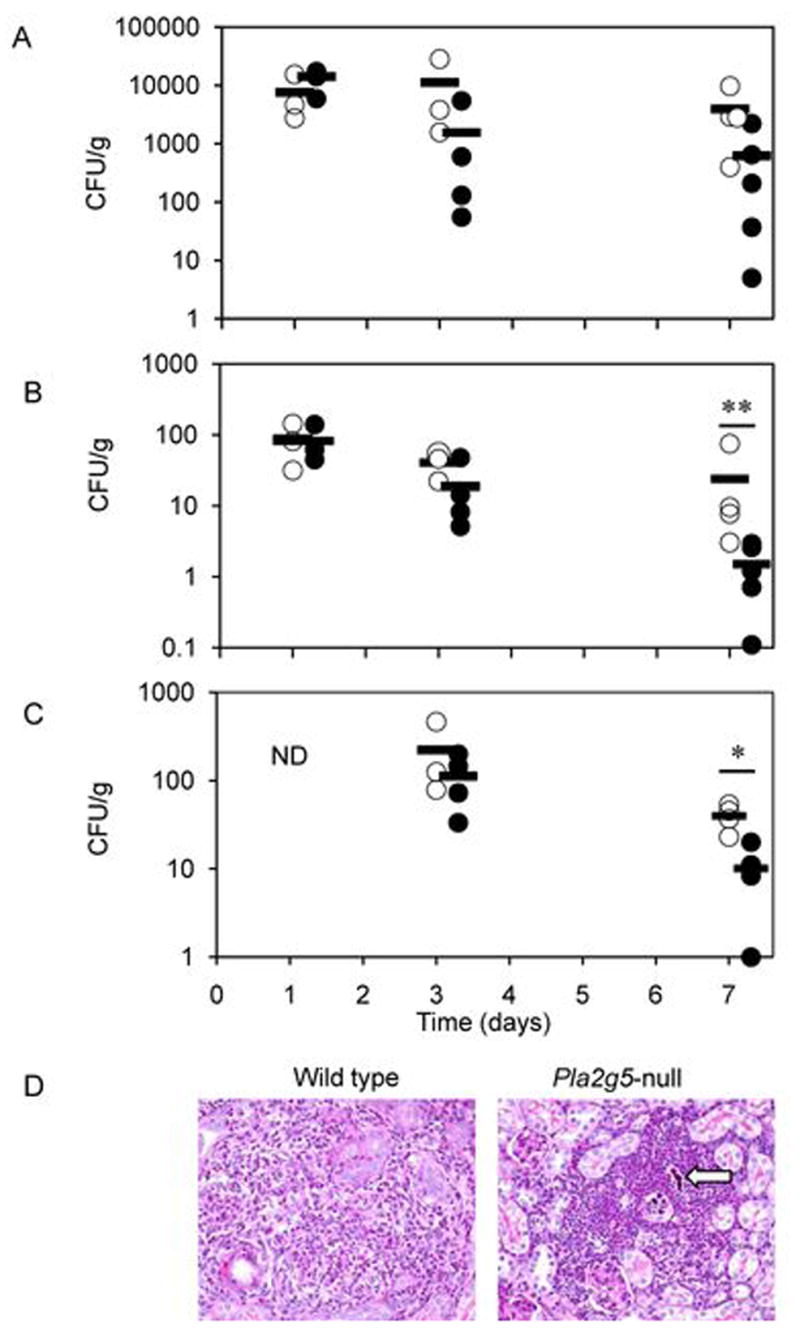
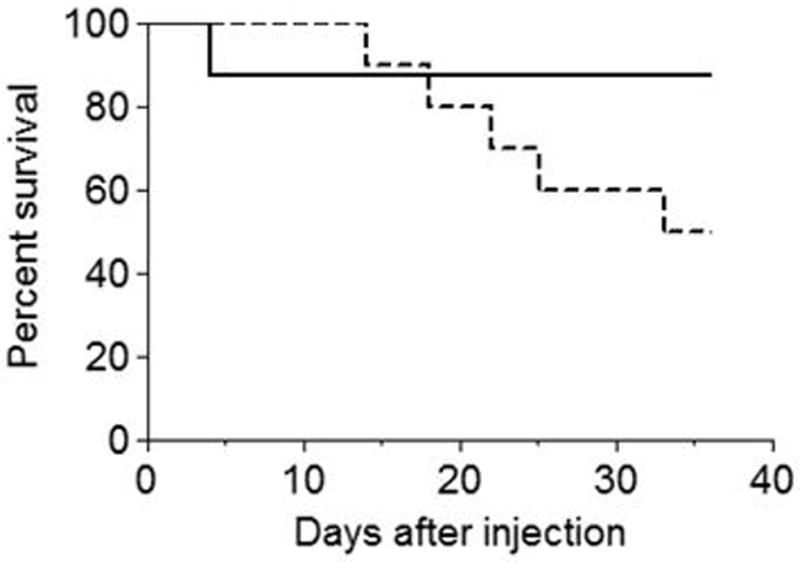
Phospholipase A(2) (PLA(2)) hydrolyzes the sn-2 position of cell membrane phospholipids to release fatty acids and lysophospholipids. We have previously reported that group V secretory PLA(2) (sPLA(2)) translocates from the Golgi and recycling endosomes of mouse peritoneal macrophages to newly formed phagosomes and regulates the phagocytosis of zymosan, suggesting a role in innate immunity. Here we report that in macrophages lacking group V sPLA(2), phagosome maturation was reduced 50-60% at early time points while the binding of zymosan was unimpaired. The ability of group V sPLA(2) to regulate phagocytosis extended to phagocytosis of IgG- and complement-opsonized sheep RBC. Moreover, macrophages lacking group V sPLA(2) had delays in phagocytosis, phagosome maturation, and killing of Candida albicans. Cytokine production and eicosanoid generation were not impaired by the lack of group V sPLA(2). Furthermore, in a model of systemic candidiasis, mice lacking group V sPLA(2) had an increased fungal burden in the kidney, liver, and spleen at day 7 postinfection and increased mortality. Thus, group V sPLA(2) regulates phagocytosis through major phagocytic receptors and contributes to the innate immune response against C. albicans by regulating phagocytosis and killing through a mechanism that is likely dependent on phagolysosome fusion.
Conflict of interest statement
Disclosures
The authors have no financial conflict of interest.
Figures
References
-
- Aderem A, Underhill DM. Mechanisms of phagocytosis in macrophages. Annu Rev Immunol. 1999;17:593–623. - PubMed
-
- Underhill DM, Ozinsky A. Phagocytosis of microbes: complexity in action. Annu Rev Immunol. 2002;20:825–852. - PubMed
-
- Taylor PR, Brown GD, Herre J, Williams DL, Willment JA, Gordon S. The role of SIGNR1 and the beta-glucan receptor (dectin-1) in the nonopsonic recognition of yeast by specific macrophages. J Immunol. 2004;172:1157–1162. - PubMed
-
- Brown GD, Gordon S. Immune recognition. A new receptor for beta-glucans. Nature. 2001;413:36–37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
