

The pattern recognition receptor CD36 is a chondrocyte hypertrophy marker associated with suppression of catabolic responses and promotion of repair responses to inflammatory stimuli
- PMID: 19342682
- PMCID: PMC2698125
- DOI: 10.4049/jimmunol.0803603
The pattern recognition receptor CD36 is a chondrocyte hypertrophy marker associated with suppression of catabolic responses and promotion of repair responses to inflammatory stimuli
Abstract
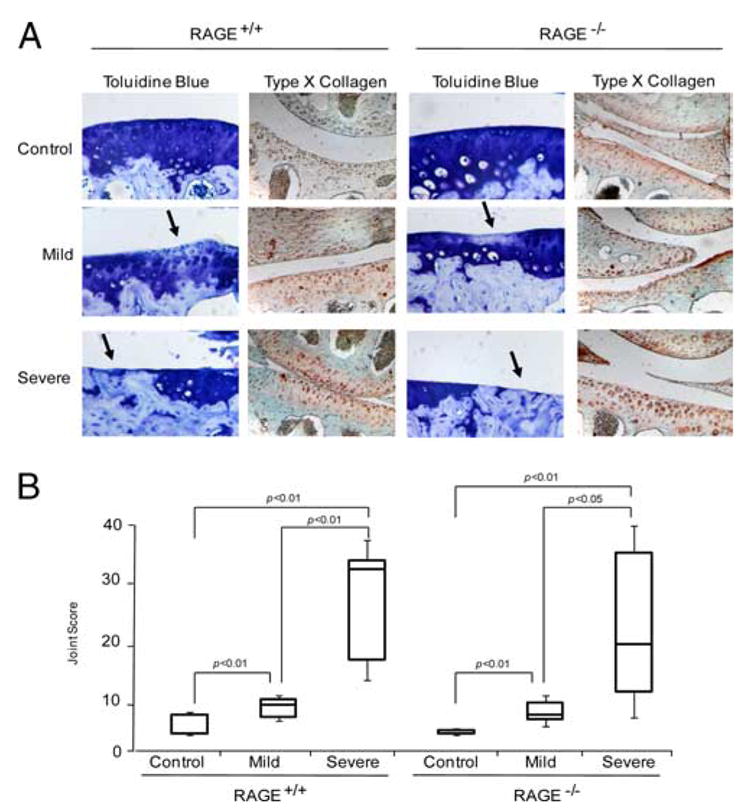
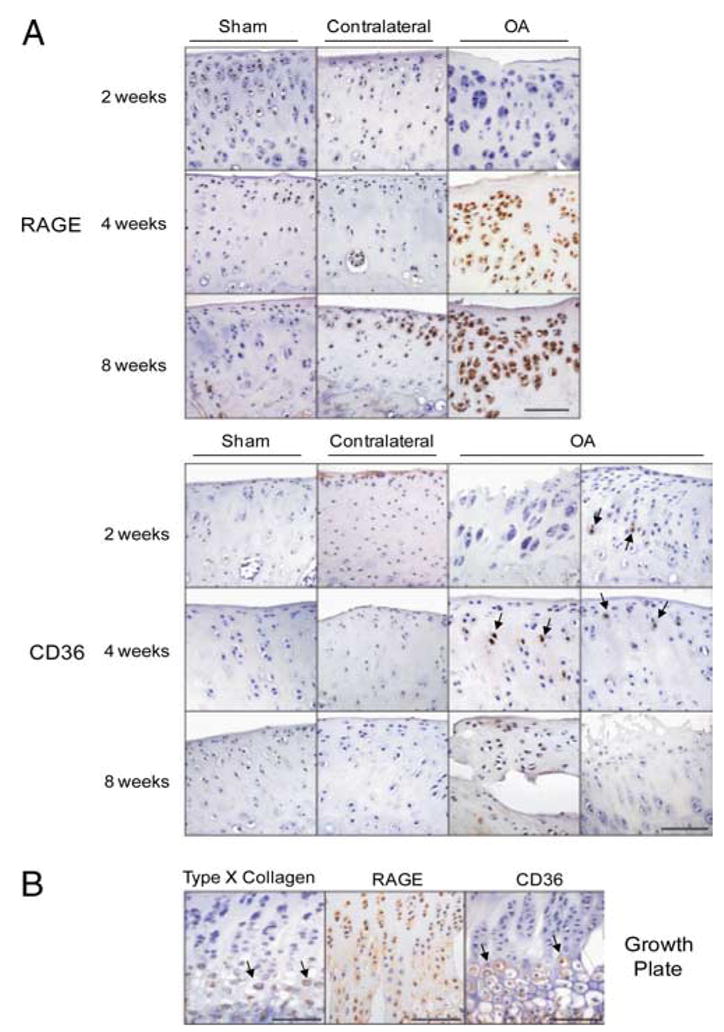
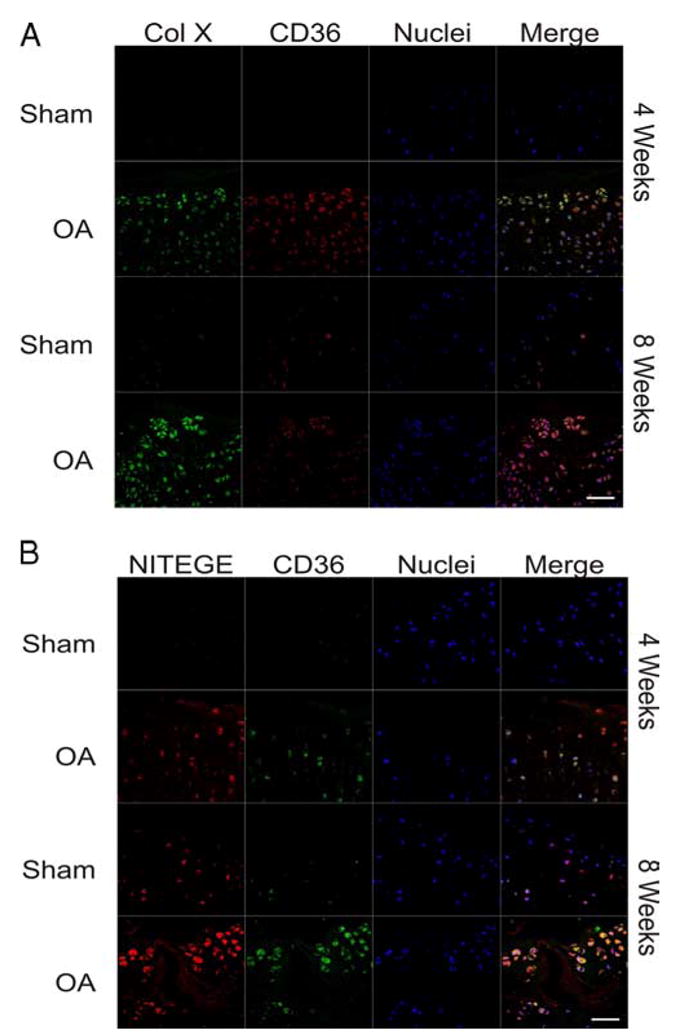
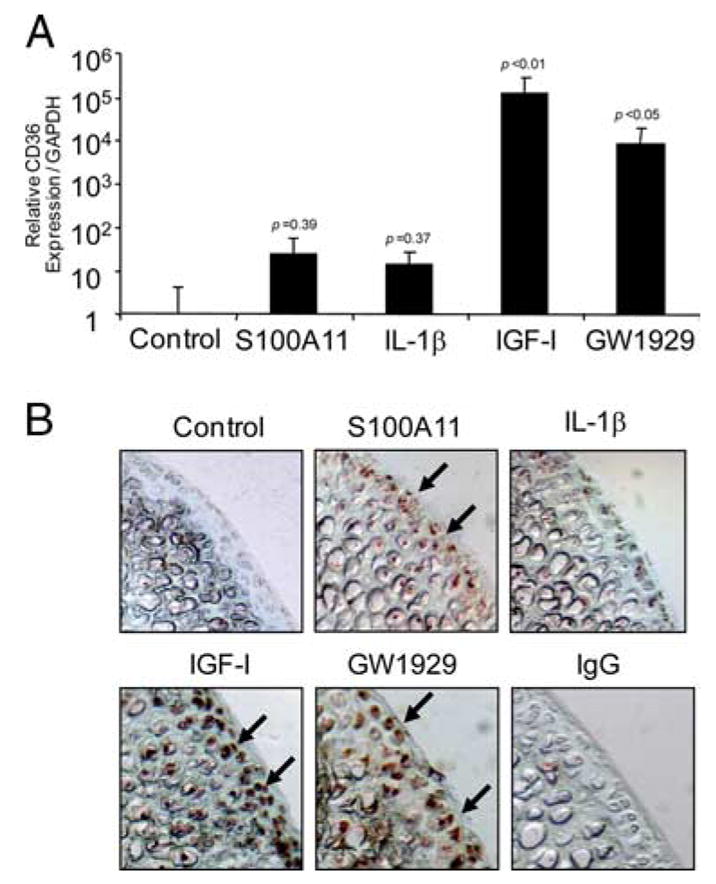
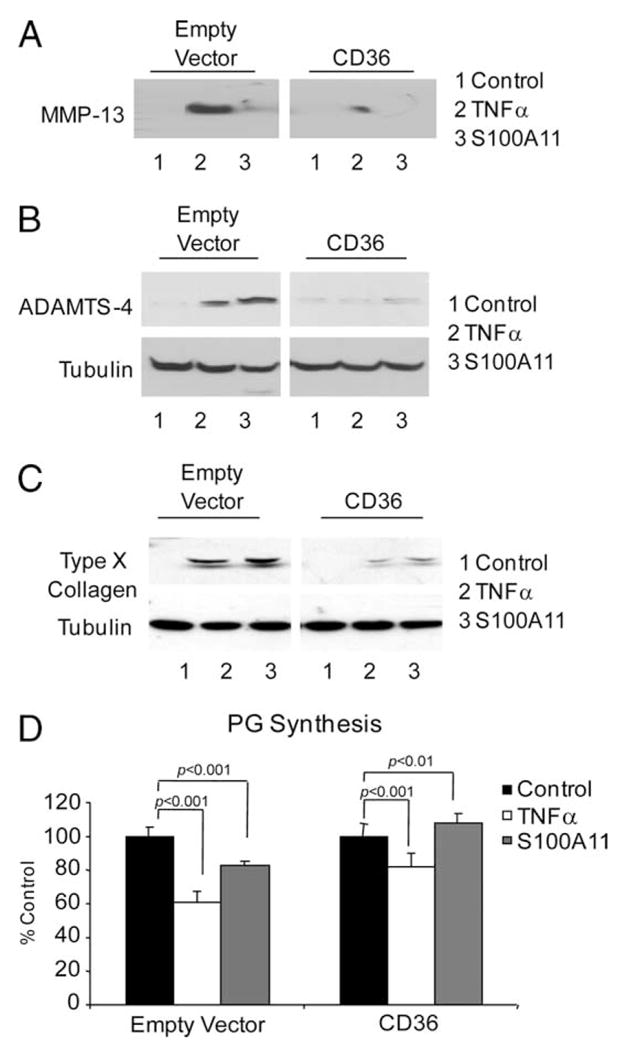
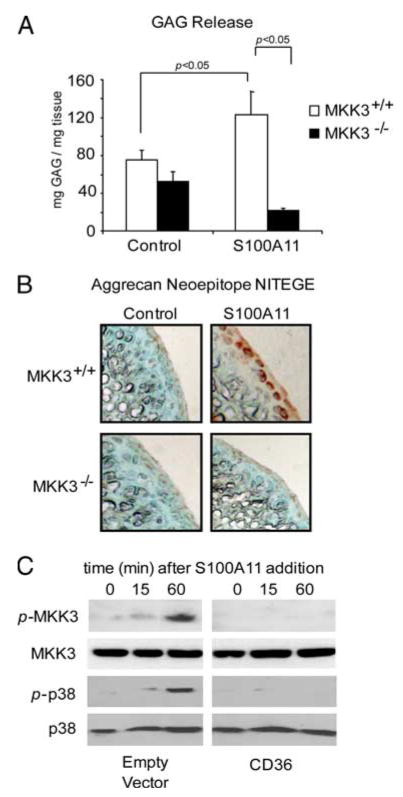
Multiple inflammatory mediators in osteoarthritis (OA) cartilage, including S100/calgranulin ligands of receptor for advanced glycation end products (RAGE), promote chondrocyte hypertrophy, a differentiation state associated with matrix catabolism. In this study, we observed that RAGE knockout was not chondroprotective in instability-induced knee OA in 8-wk-old mice. Hence, we tested the hypothesis that expression of the alternative S100/calgranulin and patterning receptor CD36, identified here as a marker of growth plate chondrocyte hypertrophy, mediates chondrocyte inflammatory and differentiation responses that promote OA. In rat knee joint destabilization-induced OA, RAGE expression was initially sparse throughout cartilage but increased diffusely by 4 wk after surgery. In contrast, CD36 expression focally increased at sites of cartilage injury and colocalized with developing chondrocyte hypertrophy and aggrecan cleavage NITEGE neoepitope formation. However, CD36 transfection in normal human knee-immortalized chondrocytes (CH-8 cells) was associated with decreased capacity of S100A11 and TNF-alpha to induce chondrocyte hypertrophy and ADAMTS-4 and matrix metalloproteinase 13 expression. S100A11 lost the capacity to inhibit proteoglycans synthesis and gained the capacity to induce proteoglycan synthesis in CD36-transfected CH-8 cells. Moreover, S100A11 required the p38 MAPK pathway kinase MKK3 to induce NITEGE development in mouse articular cartilage explants. However, CH-8 cells transfected with CD36 demonstrated decreased S100A11-induced MKK3 and p38 phosphorylation. Therefore, RAGE and CD36 patterning receptor expression were linked with opposing effects on inflammatory, procatabolic responses to S100A11 and TNF-alpha in chondrocytes.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures
References
-
- Pelletier JP, Martel-Pelletier J, Abramson SB. Osteoarthritis, an inflammatory disease: potential implication for the selection of new therapeutic targets. Arthritis Rheum. 2001;44:1237–1247. - PubMed
-
- Kobayashi M, Squires GR, Mousa A, Tanzer M, Zukor DJ, Antoniou J, Feige U, Poole AR. Role of interleukin-1 and tumor necrosis factor α in matrix degradation of human osteoarthritic cartilage. Arthritis Rheum. 2005;52:128–135. - PubMed
-
- Borzì RM, Mazzetti I, Marcu KB, Facchini A. Chemokines in cartilage degradation. Clin Orthop Relat Res. 2004;427:S53–S61. - PubMed
-
- Yammani RR, Carlson CS, Bresnick AR, Loeser RF. Increase in production of matrix metalloproteinase 13 by human articular chondrocytes due to stimulation with S100A4: role of the receptor for advanced glycation end products. Arthritis Rheum. 2006;54:2901–2911. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
