

Characterization of trans-neuronal trafficking of Cbln1
- PMID: 19344768
- PMCID: PMC2758316
- DOI: 10.1016/j.mcn.2009.03.005
Characterization of trans-neuronal trafficking of Cbln1
Abstract
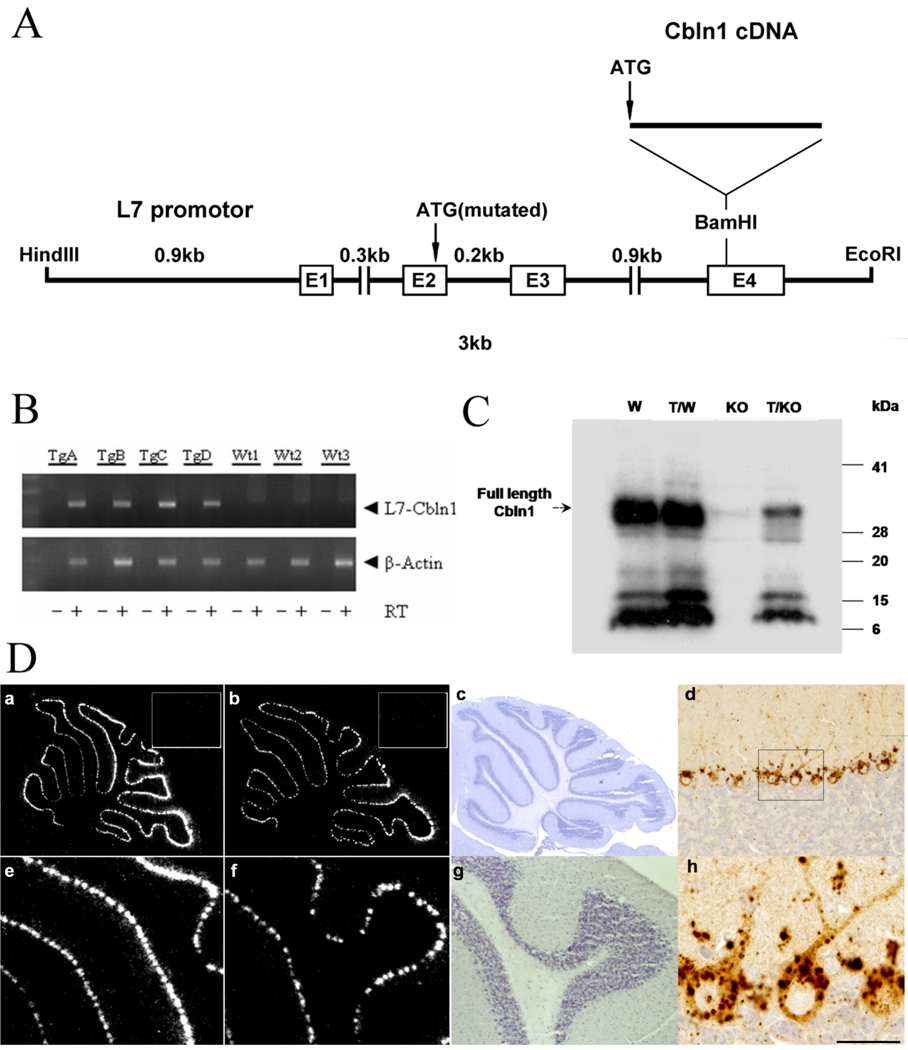
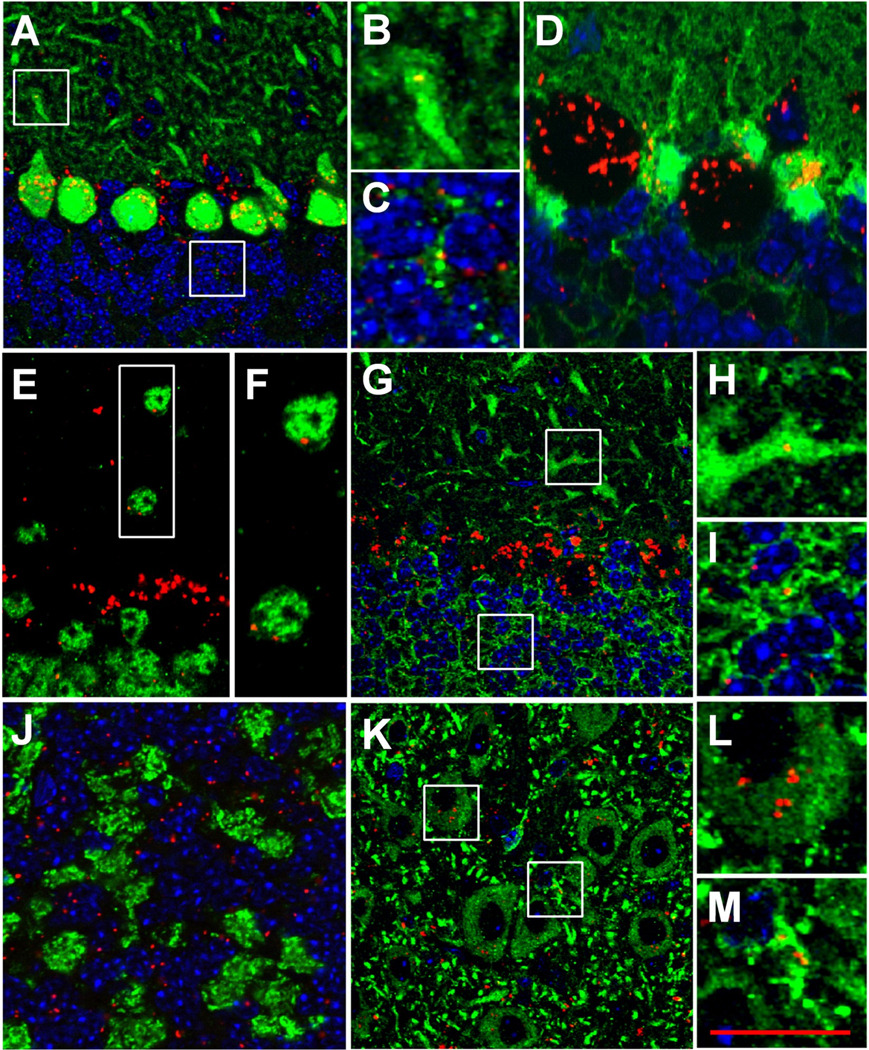
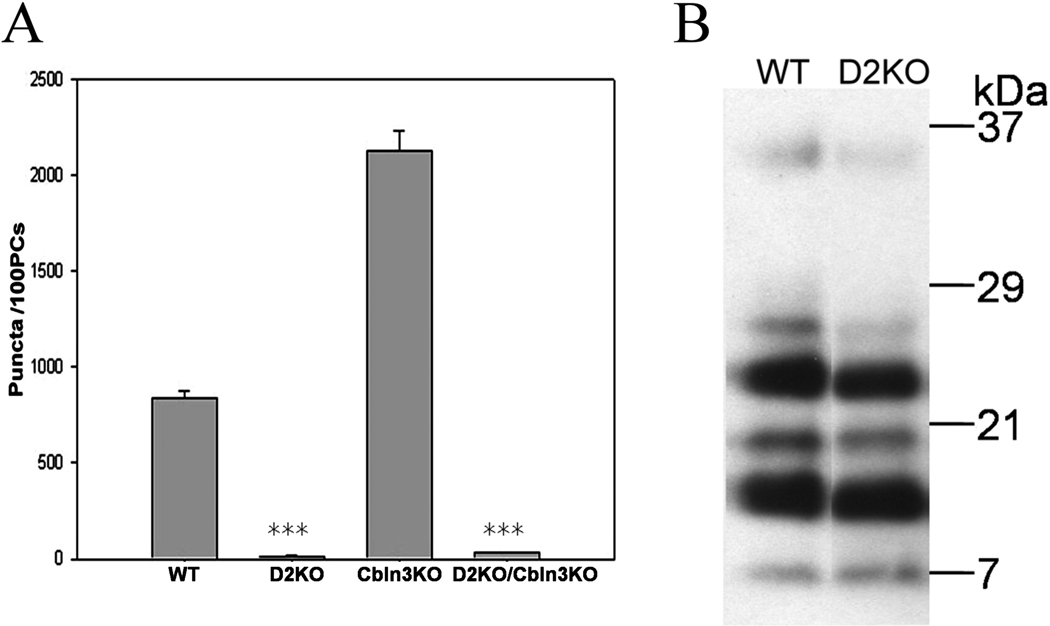
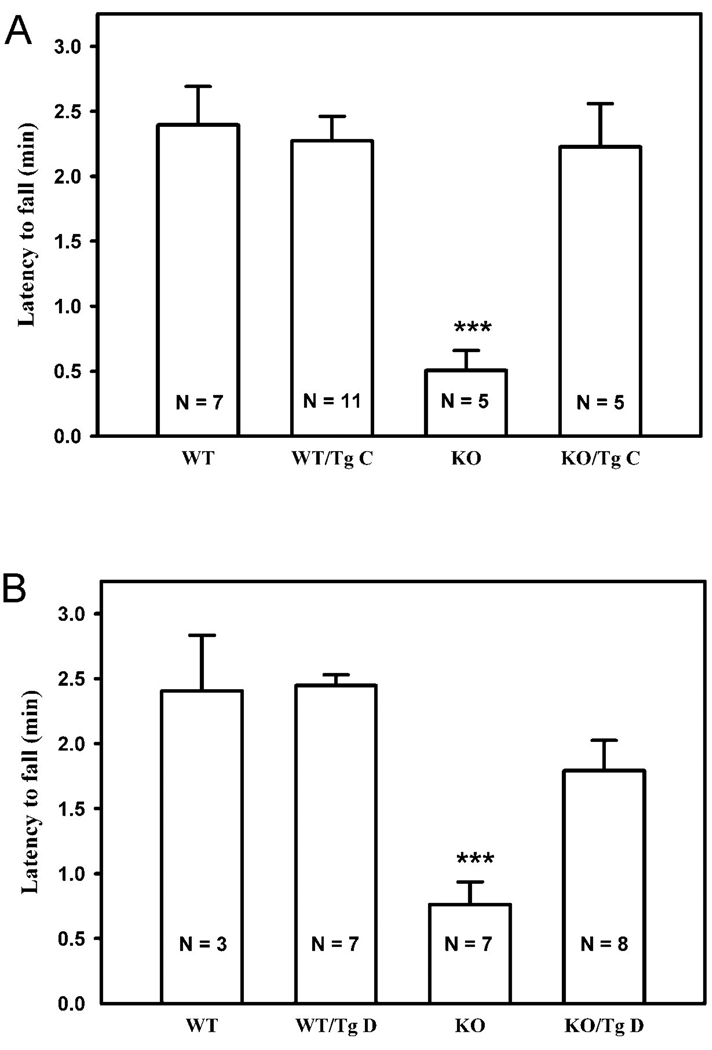
Cbln1, a glycoprotein secreted from granule cells and GluRdelta2 in the postsynaptic densities of Purkinje cells are components of an incompletely understood pathway essential for integrity and plasticity of parallel fiber-Purkinje cell synapses. We show that Cbln1 undergoes anterograde transport from granule cells to Purkinje cells and Bergmann glia, and enters the endolysosomal trafficking system, raising the possibility that Cbln1 exerts its activity on or within Purkinje cells and Bergmann glia. Cbln1 is absent in Purkinje cells and Bergmann glia of GluRdelta2-null mice, suggesting a mechanistic convergence on Cbln1 trafficking. Ectopic expression of Cbln1 in Purkinje cells of L7-cbln1 transgenic mice reveals Cbln1 undergoes anterograde and retrograde trans-neuronal trafficking even across synapses that lack GluRDelta2, indicating that it is not universally essential for Cbln1 transport. The L7-cbln1 transgene also ameliorates the locomotor deficits of cbln1-null mice, indicating that the presence and/or release of Cbln1 from the postsynaptic neuron has functional consequences.
Figures







Similar articles
-
Cbln1 accumulates and colocalizes with Cbln3 and GluRdelta2 at parallel fiber-Purkinje cell synapses in the mouse cerebellum.Eur J Neurosci. 2009 Feb;29(4):693-706. doi: 10.1111/j.1460-9568.2009.06632.x. Eur J Neurosci. 2009. PMID: 19250438 Free PMC article.
-
New (but old) molecules regulating synapse integrity and plasticity: Cbln1 and the delta2 glutamate receptor.Neuroscience. 2009 Sep 1;162(3):633-43. doi: 10.1016/j.neuroscience.2008.12.002. Epub 2008 Dec 9. Neuroscience. 2009. PMID: 19124061 Review.
-
Cbln1 downregulates the formation and function of inhibitory synapses in mouse cerebellar Purkinje cells.Eur J Neurosci. 2014 Apr;39(8):1268-80. doi: 10.1111/ejn.12487. Epub 2014 Jan 27. Eur J Neurosci. 2014. PMID: 24467251
-
Cbln1 binds to specific postsynaptic sites at parallel fiber-Purkinje cell synapses in the cerebellum.Eur J Neurosci. 2009 Feb;29(4):707-17. doi: 10.1111/j.1460-9568.2009.06639.x. Epub 2009 Feb 5. Eur J Neurosci. 2009. PMID: 19200061
-
The role of Cbln1 on Purkinje cell synapse formation.Neurosci Res. 2014 Jun;83:64-8. doi: 10.1016/j.neures.2014.01.009. Epub 2014 Mar 7. Neurosci Res. 2014. PMID: 24607546 Review.
Cited by
-
Expression of C1ql3 in Discrete Neuronal Populations Controls Efferent Synapse Numbers and Diverse Behaviors.Neuron. 2016 Sep 7;91(5):1034-1051. doi: 10.1016/j.neuron.2016.07.002. Epub 2016 Jul 28. Neuron. 2016. PMID: 27478018 Free PMC article.
-
Identification of cerebellin2 in chick and its preferential expression by subsets of developing sensory neurons and their targets in the dorsal horn.J Comp Neurol. 2010 Jul 15;518(14):2818-40. doi: 10.1002/cne.22366. J Comp Neurol. 2010. PMID: 20506477 Free PMC article.
-
Glycosylation of Cblns attenuates their receptor binding.Brain Res. 2018 Sep 1;1694:129-139. doi: 10.1016/j.brainres.2018.05.022. Epub 2018 May 18. Brain Res. 2018. PMID: 29782851 Free PMC article.
-
Comparison of Cbln1 and Cbln2 functions using transgenic and knockout mice.J Neurochem. 2012 Feb;120(4):528-40. doi: 10.1111/j.1471-4159.2011.07604.x. J Neurochem. 2012. PMID: 22117778 Free PMC article.
-
The Cbln family of proteins interact with multiple signaling pathways.J Neurochem. 2012 Jun;121(5):717-29. doi: 10.1111/j.1471-4159.2012.07648.x. Epub 2012 Feb 6. J Neurochem. 2012. PMID: 22220752 Free PMC article.
References
-
- Altar CA, DiStefano PS. Neurotrophin trafficking by anterograde transport. Trends Neurosci. 1998;21:433–437. - PubMed
-
- Bao D, Pang Z, Morgan JI. The structure and proteolytic processing of Cbln1 complexes. J. Neurochem. 2005;95:618–629. - PubMed
-
- Bellamy TC. Interactions between Purkinje neurones and Bergmann glia. Cerebellum. 2006;5:116–126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
