

A systematic survey identifies prions and illuminates sequence features of prionogenic proteins
- PMID: 19345193
- PMCID: PMC2683788
- DOI: 10.1016/j.cell.2009.02.044
A systematic survey identifies prions and illuminates sequence features of prionogenic proteins
Abstract
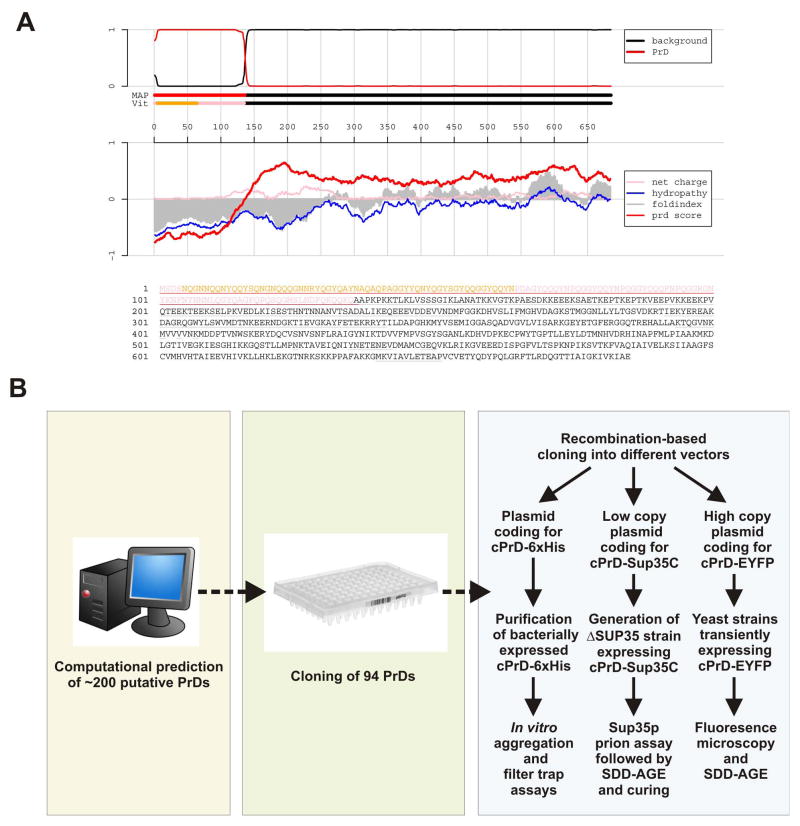
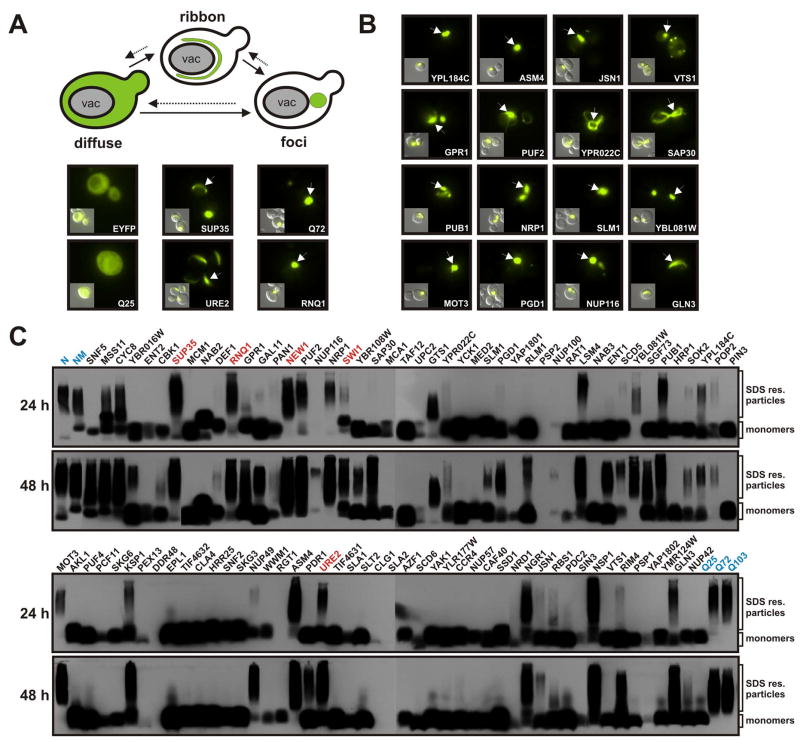
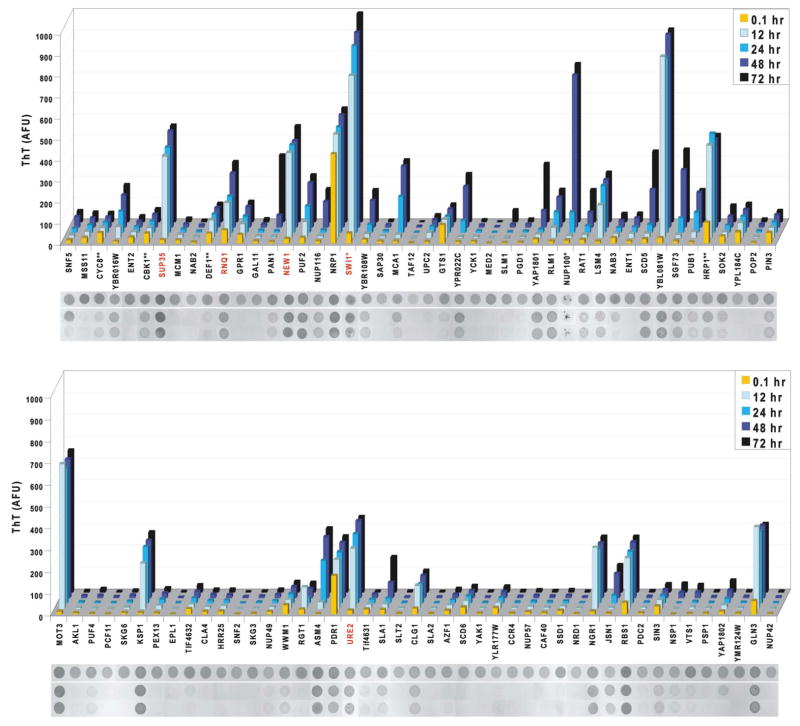
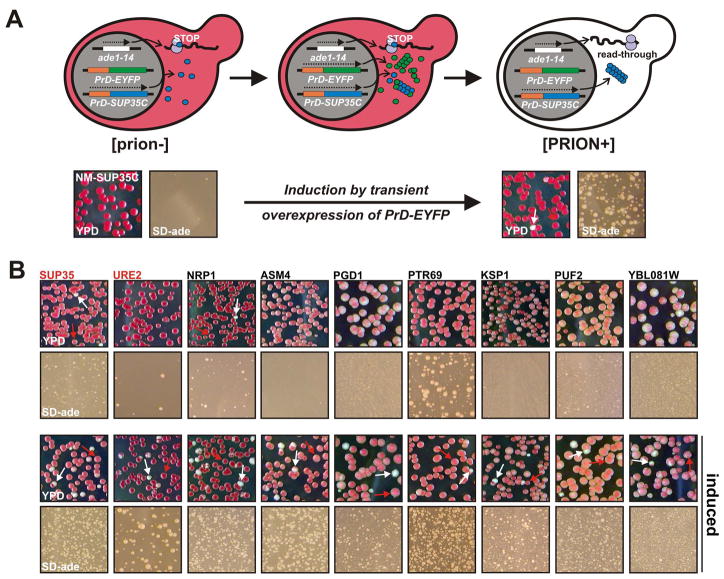
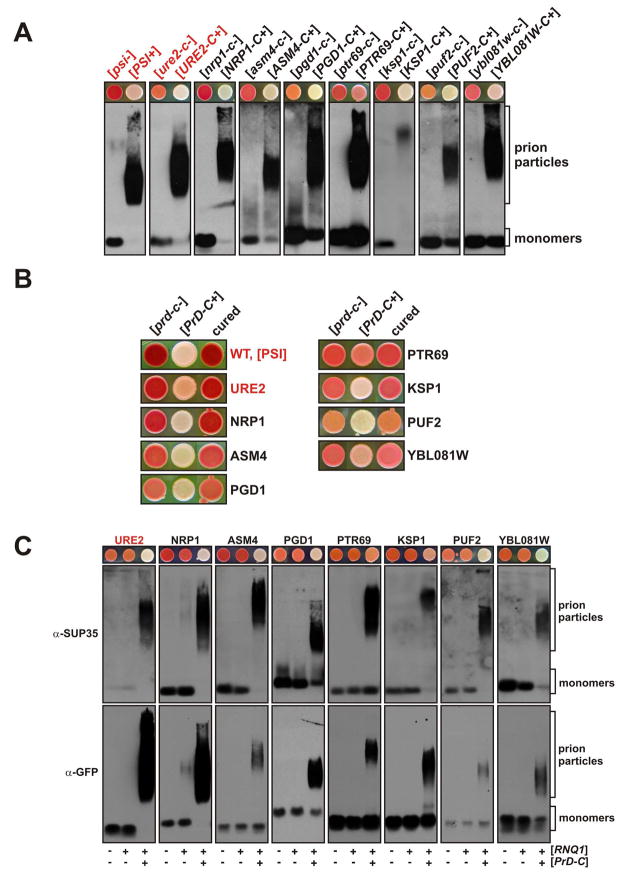
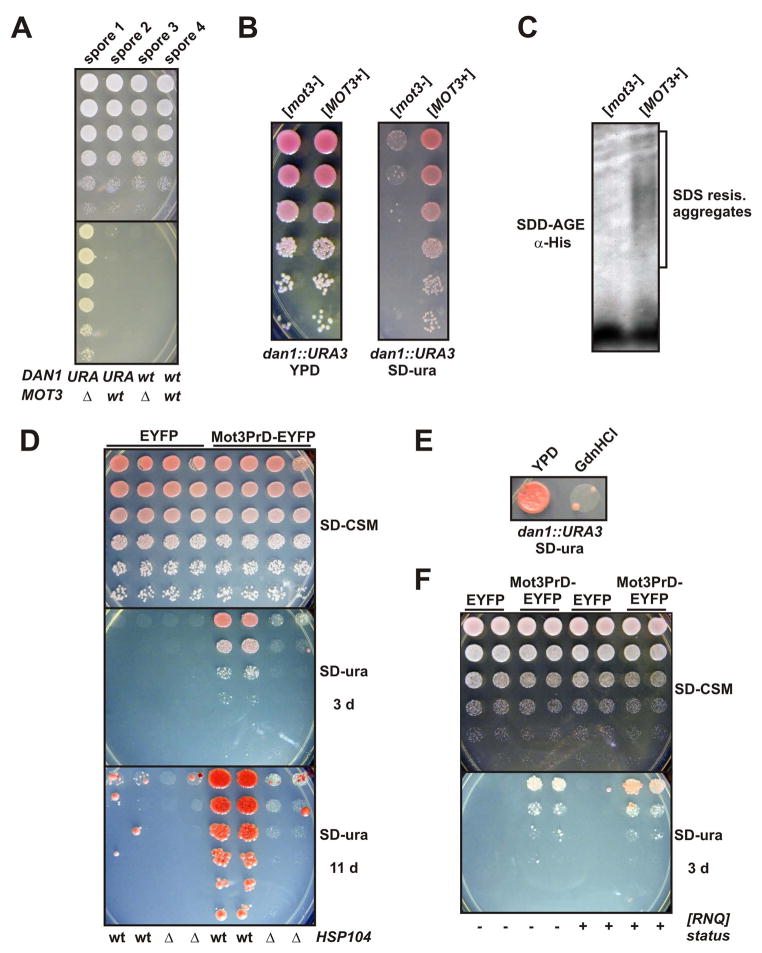
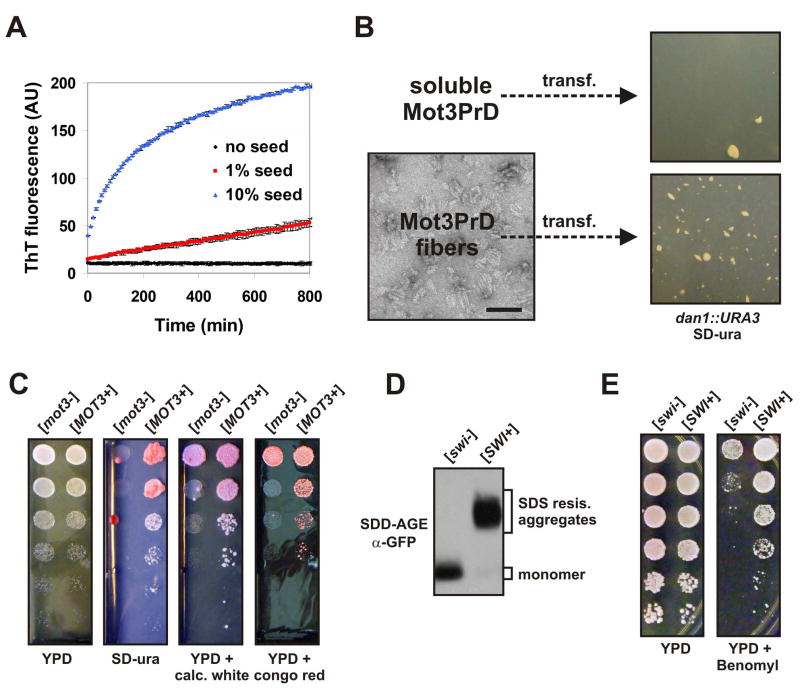
Prions are proteins that convert between structurally and functionally distinct states, one or more of which is transmissible. In yeast, this ability allows them to act as non-Mendelian elements of phenotypic inheritance. To further our understanding of prion biology, we conducted a bioinformatic proteome-wide survey for prionogenic proteins in S. cerevisiae, followed by experimental investigations of 100 prion candidates. We found an unexpected amino acid bias in aggregation-prone candidates and discovered that 19 of these could also form prions. At least one of these prion proteins, Mot3, produces a bona fide prion in its natural context that increases population-level phenotypic heterogeneity. The self-perpetuating states of these proteins present a vast source of heritable phenotypic variation that increases the adaptability of yeast populations to diverse environments.
Figures
Comment in
-
Aggregating knowledge about prions and amyloid.Cell. 2009 Apr 3;137(1):20-2. doi: 10.1016/j.cell.2009.03.030. Cell. 2009. PMID: 19345180
References
-
- Aigle M, Lacroute F. Genetical aspects of [URE3], a non-mitochondrial, cytoplasmically inherited mutation in yeast. Mol Gen Genet. 1975;136:327–335. - PubMed
-
- Bagriantsev SN, Kushnirov VV, Liebman SW. Analysis of amyloid aggregates using agarose gel electrophoresis. Methods in enzymology. 2006;412:33–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
