

Insights into a molecular switch that gates sensory neuron synapses during habituation in Aplysia
- PMID: 19345275
- PMCID: PMC4226437
- DOI: 10.1016/j.nlm.2009.03.006
Insights into a molecular switch that gates sensory neuron synapses during habituation in Aplysia
Abstract
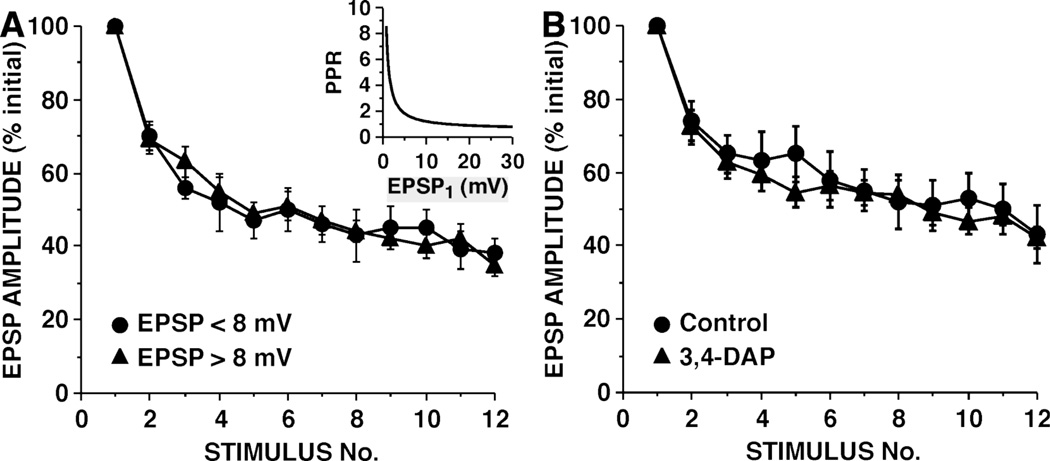
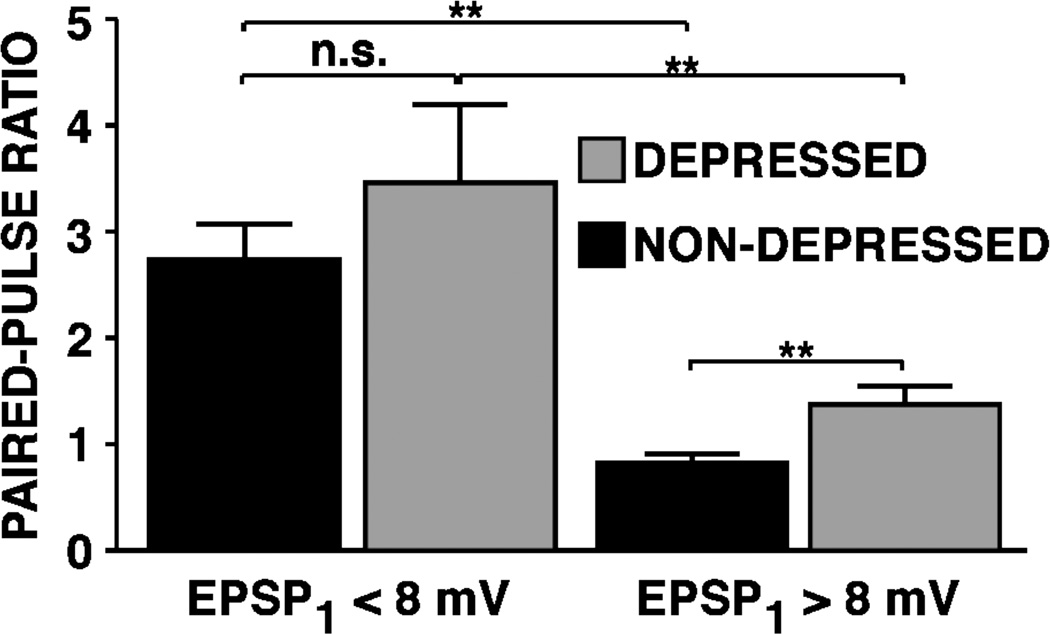
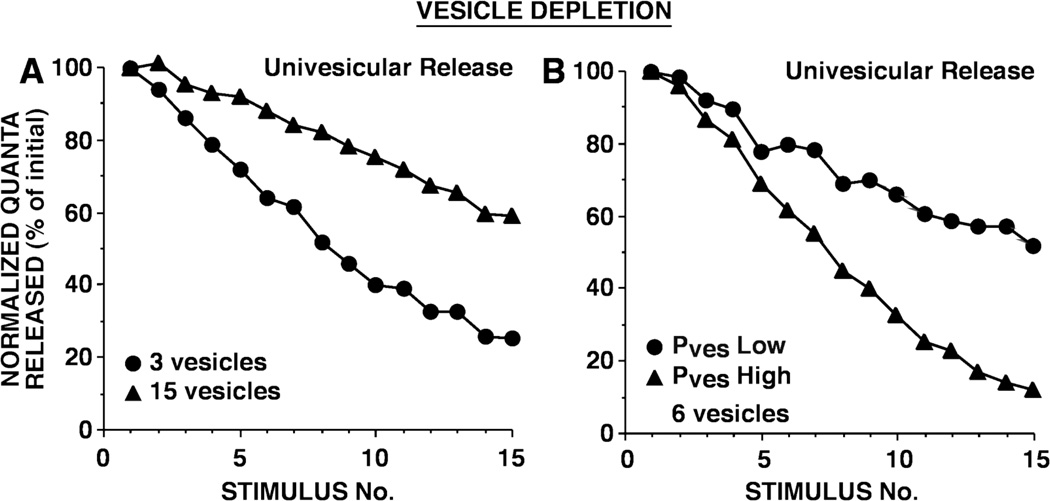
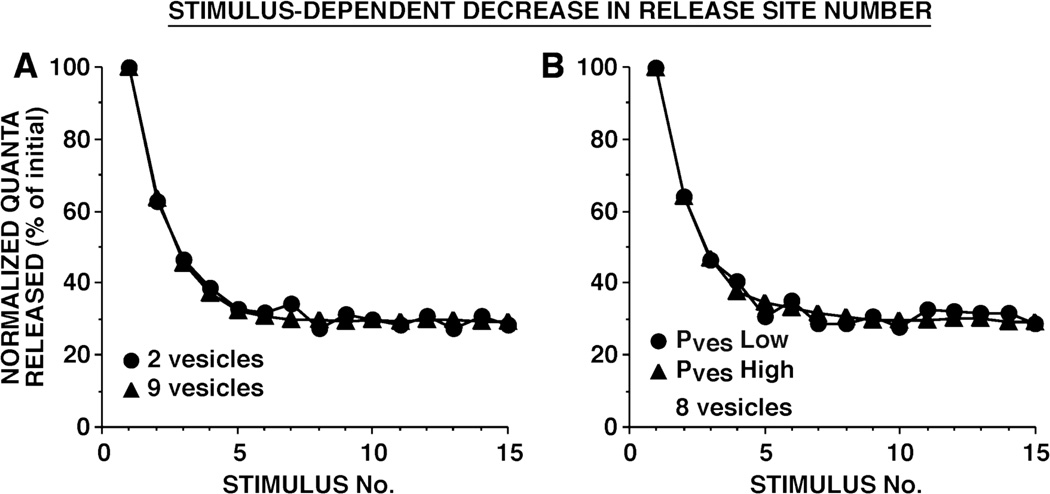
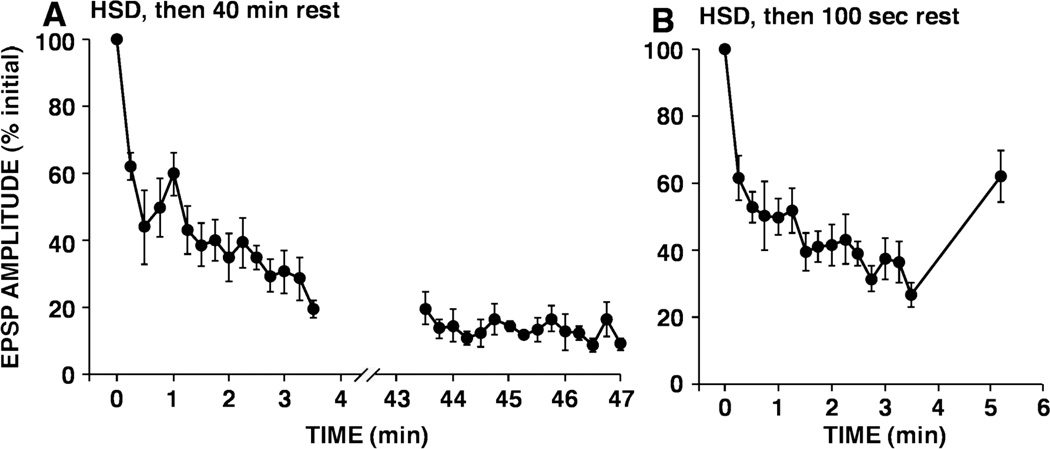
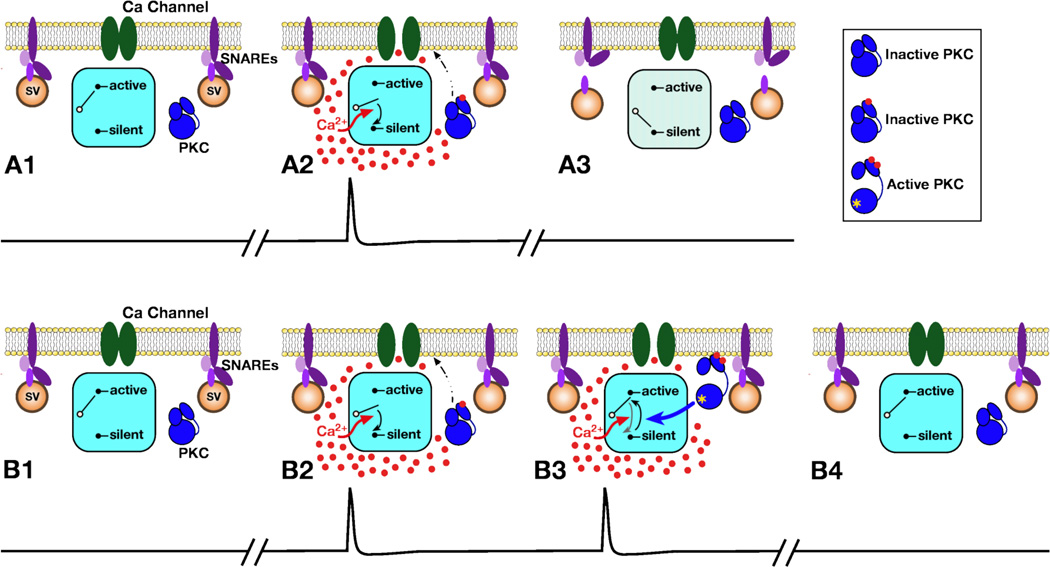
This review focuses on synaptic depression at sensory neuron-to-motor neuron synapses in the defensive withdrawal circuit of Aplysia as a model system for analysis of molecular mechanisms of sensory gating and habituation. We address the following topics: 1. Of various possible mechanisms that might underlie depression at these sensory neuron-to-motor neuron synapses in Aplysia, historically the most widely-accepted explanation has been depletion of the readily releasable pool of vesicles. Depletion is also believed to account for synaptic depression at long interstimulus intervals in a variety of other systems. 2. Multiple lines of evidence now indicate that vesicle depletion is not an important contributing mechanism to synaptic depression at Aplysia sensory neuron-to-motor neuron synapses. More generally, it appears that vesicle depletion does not contribute substantially to depression that occurs with those stimulus patterns that are typically used in studying behavioral habituation. 3. Recent evidence suggests that at these sensory neuron-to-motor neuron synapses in Aplysia, synaptic depression is mediated by an activity-dependent, but release-independent, switching of individual release sites to a silent state. This switching off of release sites is initiated by Ca2+ influx during individual action potentials. We discuss signaling proteins that may be regulated by Ca2+ during the silencing of release sites that underlies synaptic depression. 4. Bursts of 2-4 action potentials in presynaptic sensory neurons in Aplysia prevent the switching off of release sites via a mechanism called "burst-dependent protection" from synaptic depression. 5. This molecular switch may explain the sensory gating that allows animals to discriminate which stimuli are innocuous and appropriate to ignore and which stimuli are more important and should continue to elicit responses.
Figures
Similar articles
-
Simulation of synaptic depression, posttetanic potentiation, and presynaptic facilitation of synaptic potentials from sensory neurons mediating gill-withdrawal reflex in Aplysia.J Neurophysiol. 1985 Mar;53(3):652-69. doi: 10.1152/jn.1985.53.3.652. J Neurophysiol. 1985. PMID: 2580065
-
Heterosynaptic facilitation of tail sensory neuron synaptic transmission during habituation in tail-induced tail and siphon withdrawal reflexes of Aplysia.J Neurosci. 1996 Aug 15;16(16):4933-48. doi: 10.1523/JNEUROSCI.16-16-04933.1996. J Neurosci. 1996. PMID: 8756425 Free PMC article.
-
Analysis of synaptic depression contributing to habituation of gill-withdrawal reflex in Aplysia californica.J Neurophysiol. 1982 Aug;48(2):431-8. doi: 10.1152/jn.1982.48.2.431. J Neurophysiol. 1982. PMID: 6288889
-
Habituation in Aplysia: the Cheshire cat of neurobiology.Neurobiol Learn Mem. 2009 Sep;92(2):147-54. doi: 10.1016/j.nlm.2009.03.005. Epub 2009 Mar 28. Neurobiol Learn Mem. 2009. PMID: 19332142 Review.
-
Postsynaptic regulation of the development and long-term plasticity of Aplysia sensorimotor synapses in cell culture.J Neurobiol. 1994 Jun;25(6):666-93. doi: 10.1002/neu.480250608. J Neurobiol. 1994. PMID: 8071666 Review.
Cited by
-
Long-term habituation of the gill-withdrawal reflex in aplysia requires gene transcription, calcineurin and L-type voltage-gated calcium channels.Front Behav Neurosci. 2010 Nov 29;4:181. doi: 10.3389/fnbeh.2010.00181. eCollection 2010. Front Behav Neurosci. 2010. PMID: 21152260 Free PMC article.
-
Auditory cortex lesions do not disrupt habituation of HPA axis responses to repeated noise stress.Brain Res. 2012 Mar 14;1443:18-26. doi: 10.1016/j.brainres.2012.01.002. Epub 2012 Jan 13. Brain Res. 2012. PMID: 22290179 Free PMC article.
-
The CaV2α1 EF-hand F helix tyrosine, a highly conserved locus for GPCR inhibition of CaV2 channels.Sci Rep. 2018 Feb 19;8(1):3263. doi: 10.1038/s41598-018-21586-5. Sci Rep. 2018. PMID: 29459734 Free PMC article.
-
Habituation as a neural algorithm for online odor discrimination.Proc Natl Acad Sci U S A. 2020 Jun 2;117(22):12402-12410. doi: 10.1073/pnas.1915252117. Epub 2020 May 19. Proc Natl Acad Sci U S A. 2020. PMID: 32430320 Free PMC article.
-
Habituation of reflexive and motivated behavior in mice with deficient BK channel function.Front Integr Neurosci. 2013 Nov 19;7:79. doi: 10.3389/fnint.2013.00079. eCollection 2013. Front Integr Neurosci. 2013. PMID: 24312024 Free PMC article.
References
-
- Antonov I, Antonova I, Kandel ER, Hawkins RD. Activity-dependent presynaptic facilitation and hebbian LTP are both required and interact during classical conditioning in Aplysia. Neuron. 2003;37:135–147. - PubMed
-
- Antzoulatos EG, Cleary LJ, Eskin A, Baxter DA, Byrne JH. Desensitization of postsynaptic glutamate receptors contributes to high-frequency homosynaptic depression of aplysia sensorimotor connections. Learn Mem. 2003;10:309–313. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
