

Globular tetramers of beta(2)-microglobulin assemble into elaborate amyloid fibrils
- PMID: 19345691
- PMCID: PMC2726924
- DOI: 10.1016/j.jmb.2009.03.066
Globular tetramers of beta(2)-microglobulin assemble into elaborate amyloid fibrils
Abstract
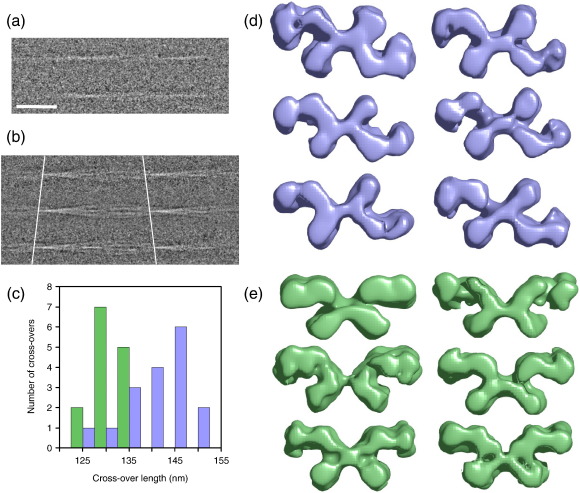
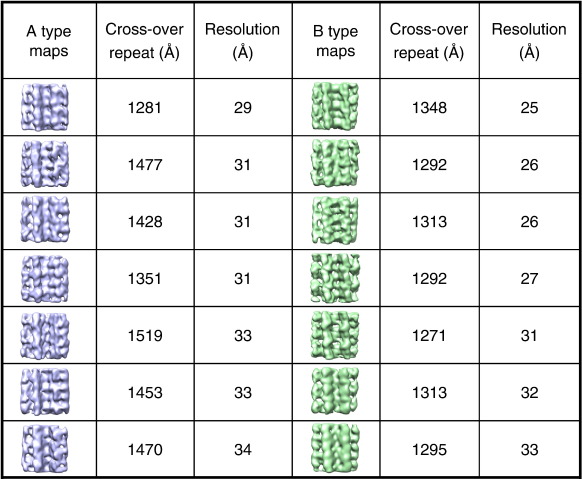
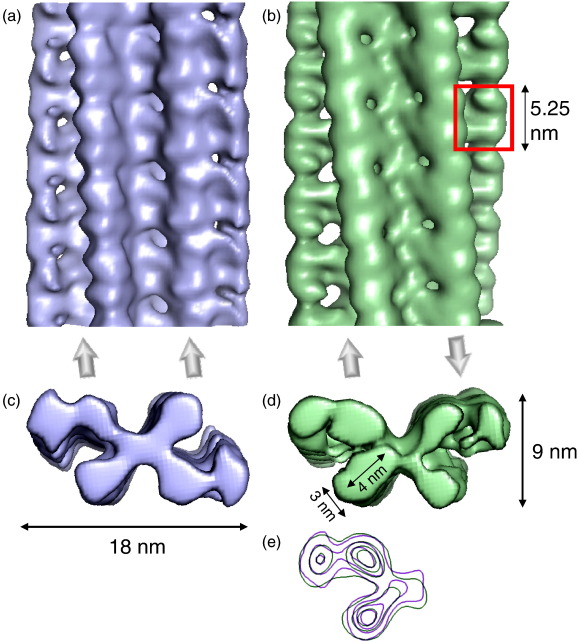
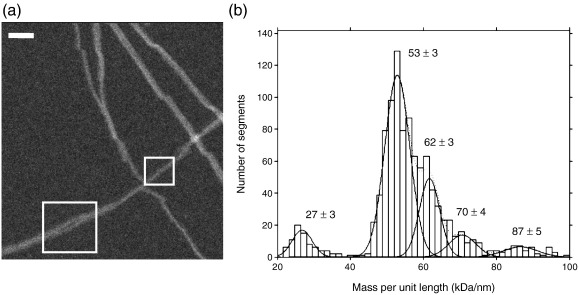
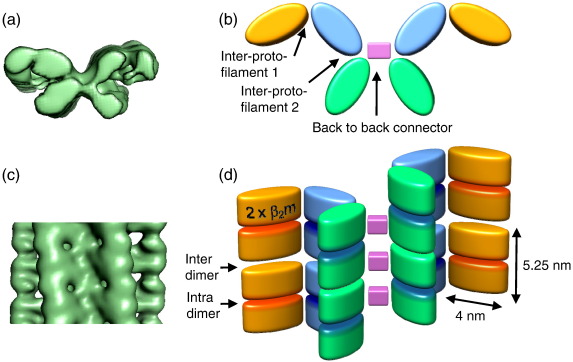
Amyloid fibrils are ordered polymers in which constituent polypeptides adopt a non-native fold. Despite their importance in degenerative human diseases, the overall structure of amyloid fibrils remains unknown. High-resolution studies of model peptide assemblies have identified residues forming cross-beta-strands and have revealed some details of local beta-strand packing. However, little is known about the assembly contacts that define the fibril architecture. Here we present a set of three-dimensional structures of amyloid fibrils formed from full-length beta(2)-microglobulin, a 99-residue protein involved in clinical amyloidosis. Our cryo-electron microscopy maps reveal a hierarchical fibril structure built from tetrameric units of globular density, with at least three different subunit interfaces in this homopolymeric assembly. These findings suggest a more complex superstructure for amyloid than hitherto suspected and prompt a re-evaluation of the defining features of the amyloid fold.
Figures

Similar articles
-
The structure of a β2-microglobulin fibril suggests a molecular basis for its amyloid polymorphism.Nat Commun. 2018 Oct 30;9(1):4517. doi: 10.1038/s41467-018-06761-6. Nat Commun. 2018. PMID: 30375379 Free PMC article.
-
Secondary structure in the core of amyloid fibrils formed from human β₂m and its truncated variant ΔN6.J Am Chem Soc. 2014 Apr 30;136(17):6313-25. doi: 10.1021/ja4126092. Epub 2014 Apr 16. J Am Chem Soc. 2014. PMID: 24679070 Free PMC article.
-
Beta2-microglobulin amyloid fragment organization and morphology and its comparison to Abeta suggests that amyloid aggregation pathways are sequence specific.Biochemistry. 2008 Feb 26;47(8):2497-509. doi: 10.1021/bi7019194. Epub 2008 Jan 24. Biochemistry. 2008. PMID: 18215070
-
Structural stability of amyloid fibrils of beta(2)-microglobulin in comparison with its native fold.Biochim Biophys Acta. 2005 Nov 10;1753(1):64-75. doi: 10.1016/j.bbapap.2005.08.002. Epub 2005 Aug 24. Biochim Biophys Acta. 2005. PMID: 16213801 Review.
-
Fibril modelling by sequence and structure conservation analysis combined with protein docking techniques: beta(2)-microglobulin amyloidosis.Biochim Biophys Acta. 2005 Nov 10;1753(1):121-30. doi: 10.1016/j.bbapap.2005.07.012. Epub 2005 Aug 2. Biochim Biophys Acta. 2005. PMID: 16107326 Review.
Cited by
-
The Early Phase of β2-Microglobulin Aggregation: Perspectives From Molecular Simulations.Front Mol Biosci. 2020 Sep 29;7:578433. doi: 10.3389/fmolb.2020.578433. eCollection 2020. Front Mol Biosci. 2020. PMID: 33134317 Free PMC article. Review.
-
A covalent homodimer probing early oligomers along amyloid aggregation.Sci Rep. 2015 Sep 30;5:14651. doi: 10.1038/srep14651. Sci Rep. 2015. PMID: 26420657 Free PMC article.
-
Optical trapping with high forces reveals unexpected behaviors of prion fibrils.Nat Struct Mol Biol. 2010 Dec;17(12):1422-30. doi: 10.1038/nsmb.1954. Epub 2010 Nov 28. Nat Struct Mol Biol. 2010. PMID: 21113168 Free PMC article.
-
The amyloid state of proteins in human diseases.Cell. 2012 Mar 16;148(6):1188-203. doi: 10.1016/j.cell.2012.02.022. Cell. 2012. PMID: 22424229 Free PMC article. Review.
-
Structural insights into the pre-amyloid tetramer of β-2-microglobulin from covalent labeling and mass spectrometry.Biochemistry. 2011 Aug 9;50(31):6711-22. doi: 10.1021/bi2004894. Epub 2011 Jul 8. Biochemistry. 2011. PMID: 21718071 Free PMC article.
References
-
- Cohen A.S., Calkins E. Electron microscopic observations on a fibrous component in amyloid of diverse origins. Nature. 1959;183:1202–1203. - PubMed
-
- Pepys M.B. Amyloidosis. Annu. Rev. Med. 2006;57:223–241. - PubMed
-
- Fowler D.M., Koulov A.V., Balch W.E., Kelly J.W. Functional amyloid — from bacteria to humans. Trends Biochem. Sci. 2007;32:217–224. - PubMed
-
- Chiti F., Dobson C.M. Protein misfolding, functional amyloid, and human disease. Annu. Rev. Biochem. 2006;75:333–366. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
