

The contribution of Ca2+ signaling and Ca2+ sensitivity to the regulation of airway smooth muscle contraction is different in rats and mice
- PMID: 19346434
- PMCID: PMC2692797
- DOI: 10.1152/ajplung.90288.2008
The contribution of Ca2+ signaling and Ca2+ sensitivity to the regulation of airway smooth muscle contraction is different in rats and mice
Abstract
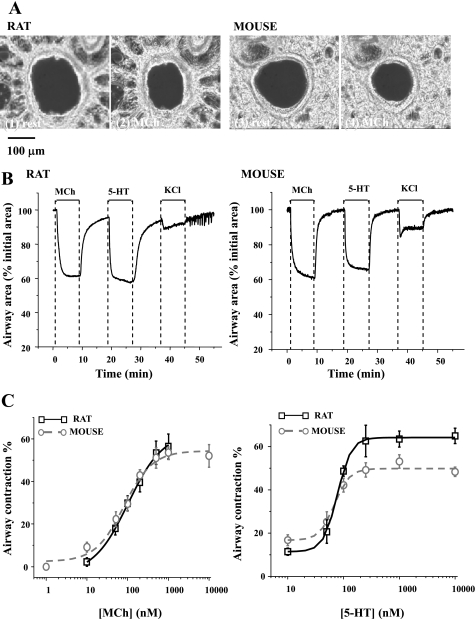
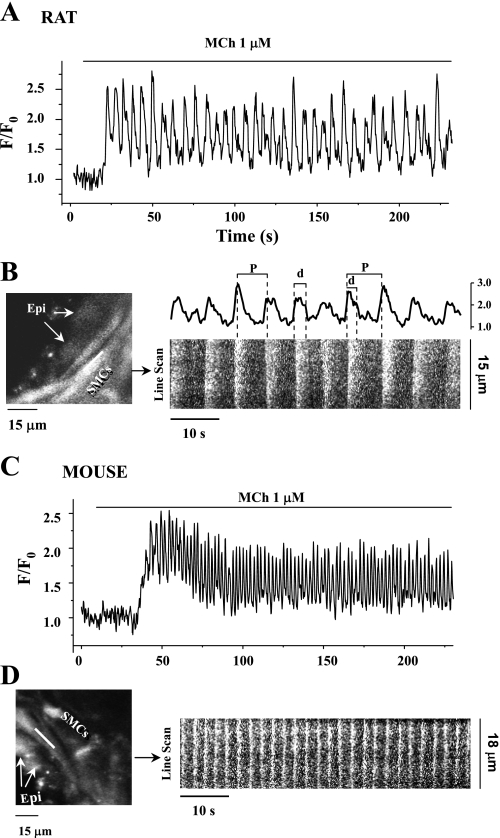
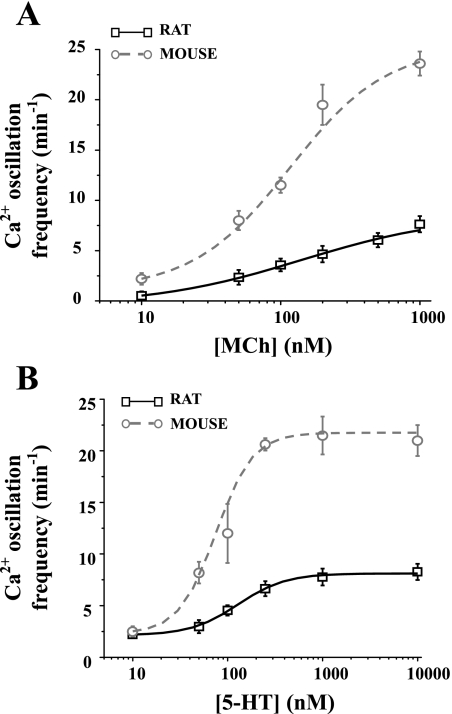
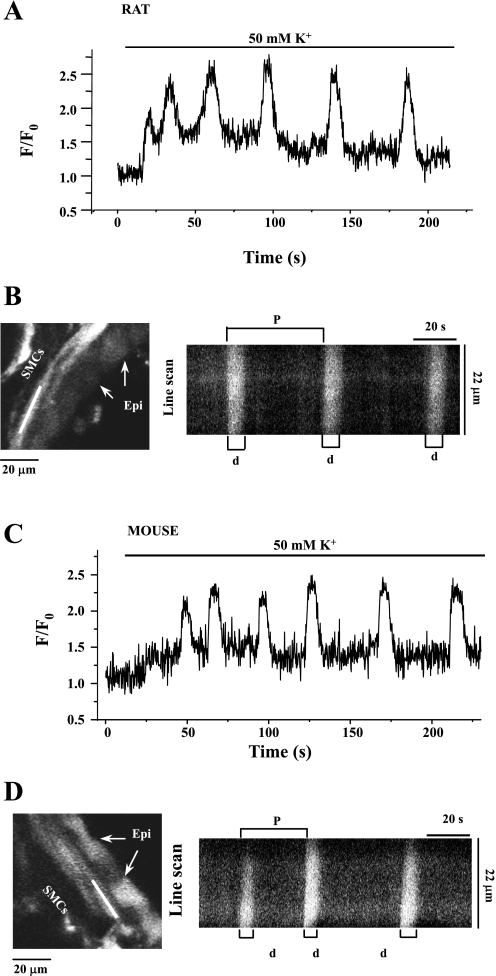
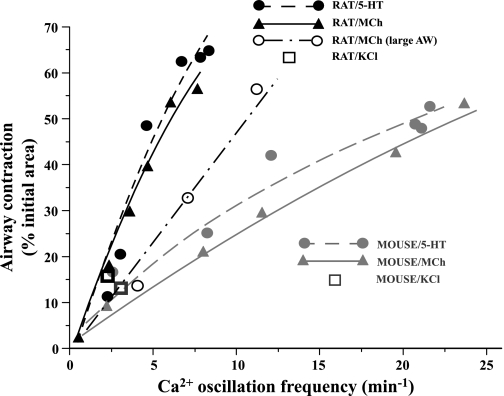
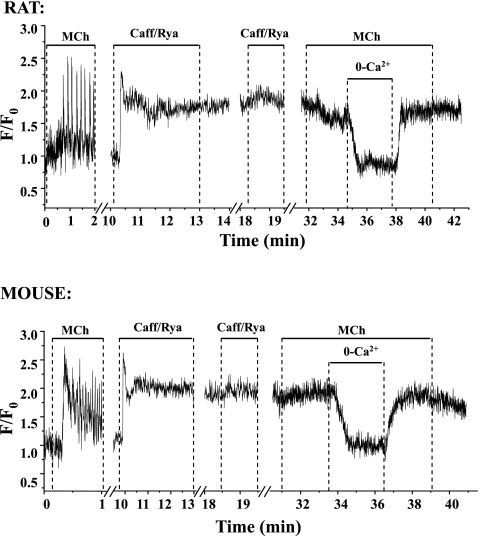
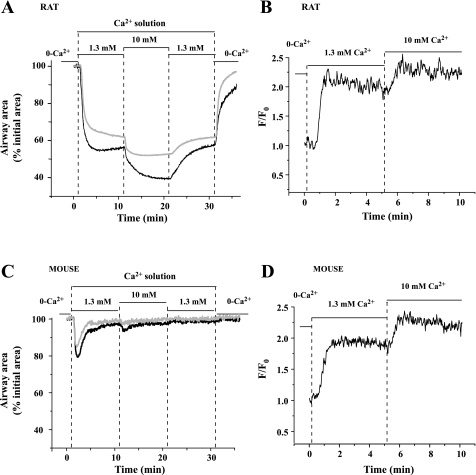
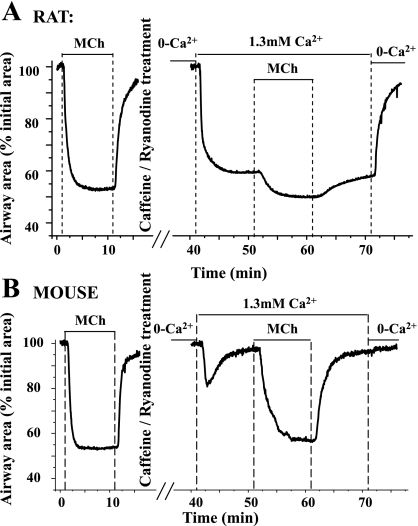
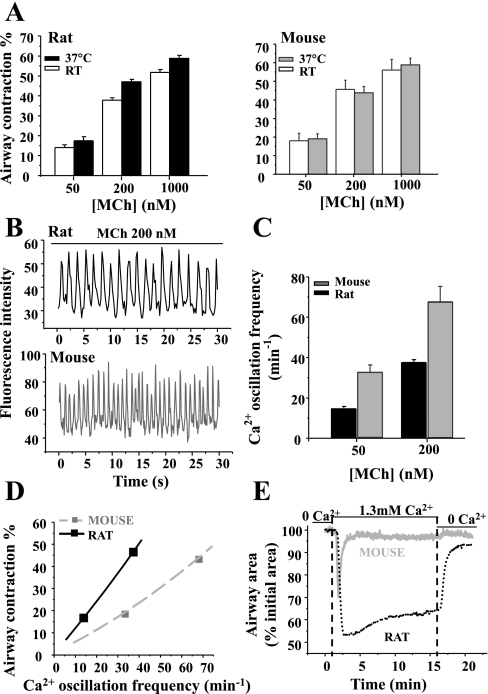
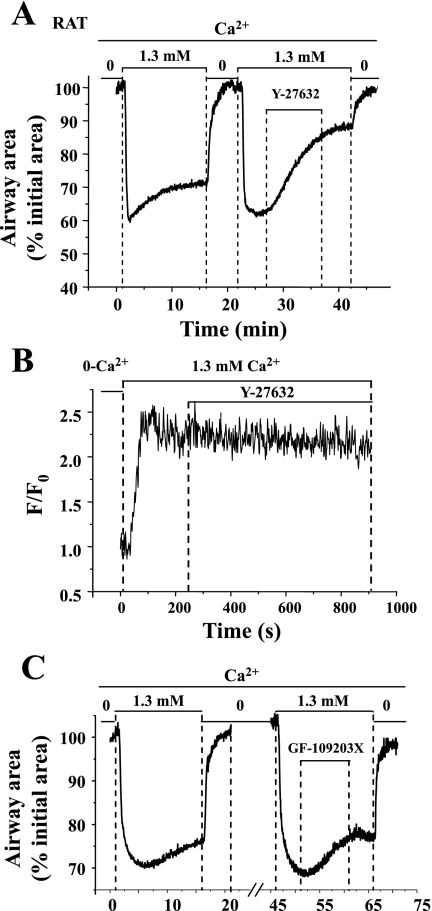
To determine the relative contributions of Ca(2+) signaling and Ca(2+) sensitivity to the contractility of airway smooth muscle cells (SMCs), we compared the contractile responses of mouse and rat airways with the lung slice technique. Airway contraction was measured by monitoring changes in airway lumen area with phase-contrast microscopy, whereas changes in intracellular calcium concentration ([Ca(2+)](i)) of the SMCs were recorded with laser scanning microscopy. In mice and rats, methacholine (MCh) or serotonin induced concentration-dependent airway contraction and Ca(2+) oscillations in the SMCs. However, rat airways demonstrated greater contraction compared with mice, in response to agonist-induced Ca(2+) oscillations of a similar frequency. Because this indicates that rat airway SMCs have a higher Ca(2+) sensitivity compared with mice, we examined Ca(2+) sensitivity with Ca(2+)-permeabilized airway SMCs in which the [Ca(2+)](i) was experimentally controlled. In the absence of agonists, high [Ca(2+)](i) induced a sustained contraction in rat airways but only a transient contraction in mouse airways. This sustained contraction of rat airways was relaxed by Y-23672, a Rho kinase inhibitor, but not affected by GF-109203X, a PKC inhibitor. The subsequent exposure of Ca(2+)-permeabilized airway SMCs, with high [Ca(2+)](i), to MCh elicited a further contraction of rat airways and initiated a sustained contraction of mouse airways, without changing the [Ca(2+)](i) of the SMCs. Collectively, these results indicate that airway SMCs of rats have a substantially higher innate Ca(2+) sensitivity than mice and that this strongly influences the transduction of the frequency of Ca(2+) oscillations into the contractility of airway SMCs.
Figures
References
-
- Bai Y, Sanderson MJ. Modulation of the Ca2+ sensitivity of airway smooth muscle cells in murine lung slices. Am J Physiol Lung Cell Mol Physiol 291: L208–L221, 2006. - PubMed
-
- Bergner A, Sanderson MJ. Airway contractility and smooth muscle Ca2+ signaling in lung slices from different mouse strains. J Appl Physiol 95: 1325–1332; discussion 1314, 2003. - PubMed
-
- Björck T, Gustafsson LE, Dahlén SE. Isolated bronchi from asthmatics are hyperresponsive to adenosine, which apparently acts indirectly by liberation of leukotrienes and histamine. Am Rev Respir Dis 145: 1087–1091, 1992. - PubMed
-
- Black JL Pharmacology of airway smooth muscle in chronic obstructive pulmonary disease and in asthma. Am Rev Respir Dis 143: 1177–1181, 1991. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous