

Single amino acid substitution in Plasmodium yoelii erythrocyte ligand determines its localization and controls parasite virulence
- PMID: 19346470
- PMCID: PMC2678433
- DOI: 10.1073/pnas.0811313106
Single amino acid substitution in Plasmodium yoelii erythrocyte ligand determines its localization and controls parasite virulence
Abstract
The major virulence determinant of the rodent malaria parasite, Plasmodium yoelii, has remained unresolved since the discovery of the lethal line in the 1970s. Because virulence in this parasite correlates with the ability to invade different types of erythrocytes, we evaluated the potential role of the parasite erythrocyte binding ligand, PyEBL. We found 1 amino acid substitution in a domain responsible for intracellular trafficking between the lethal and nonlethal parasite lines and, furthermore, that the intracellular localization of PyEBL was distinct between these lines. Genetic modification showed that this substitution was responsible not only for PyEBL localization but also the erythrocyte-type invasion preference of the parasite and subsequently its virulence in mice. This previously unrecognized mechanism for altering an invasion phenotype indicates that subtle alterations of a malaria parasite ligand can dramatically affect host-pathogen interactions and malaria virulence.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

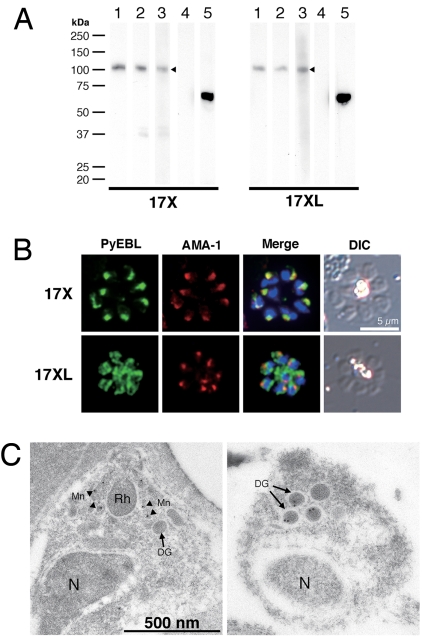
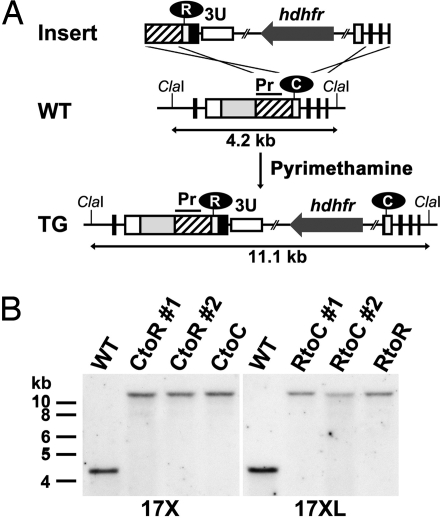
References
-
- Landau I, Gautret P. In: Malaria: Parasite Biology, Pathogenesis, and Protection. Sherman IW, editor. Washington, DC: American Society for Microbiology; 1998. pp. 401–417.
-
- Yoeli M, Hargreaves B, Carter R, Walliker D. Sudden increase in virulence in a strain of Plasmodium berghei yoelii. Ann Trop Med Parasitol. 1975;69:173–178. - PubMed
-
- Miller LH, Mason SJ, Clyde DF, McGinniss MH. The resistance factor to Plasmodium vivax in blacks. The Duffy-blood-group genotype, FyFy. N Engl J Med. 1976;295:302–304. - PubMed
-
- Wertheimer SP, Barnwell JW. Plasmodium vivax interaction with the human Duffy blood group glycoprotein: Identification of a parasite receptor-like protein. Exp Parasitol. 1989;69:340–350. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
