

Clustering of InsP3 receptors by InsP3 retunes their regulation by InsP3 and Ca2+
- PMID: 19348050
- PMCID: PMC2702691
- DOI: 10.1038/nature07763
Clustering of InsP3 receptors by InsP3 retunes their regulation by InsP3 and Ca2+
Abstract
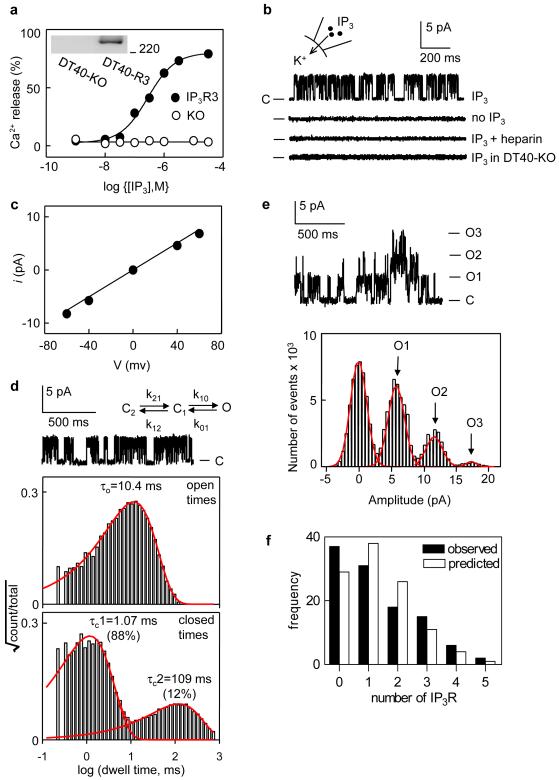
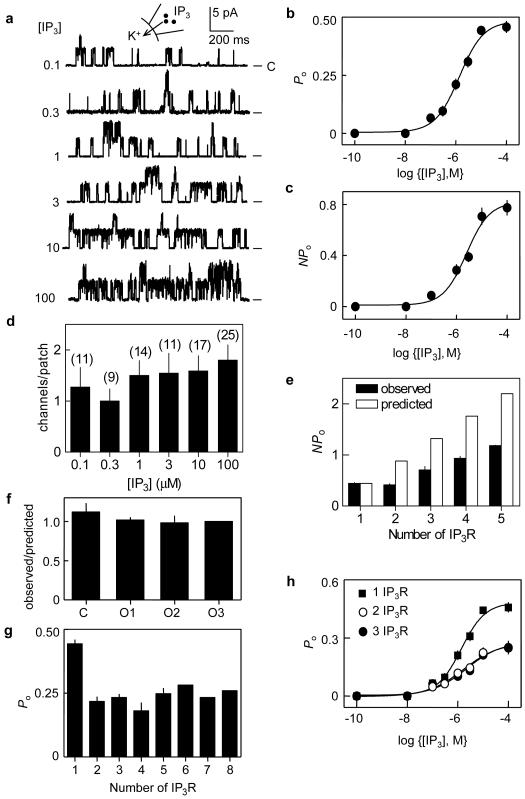
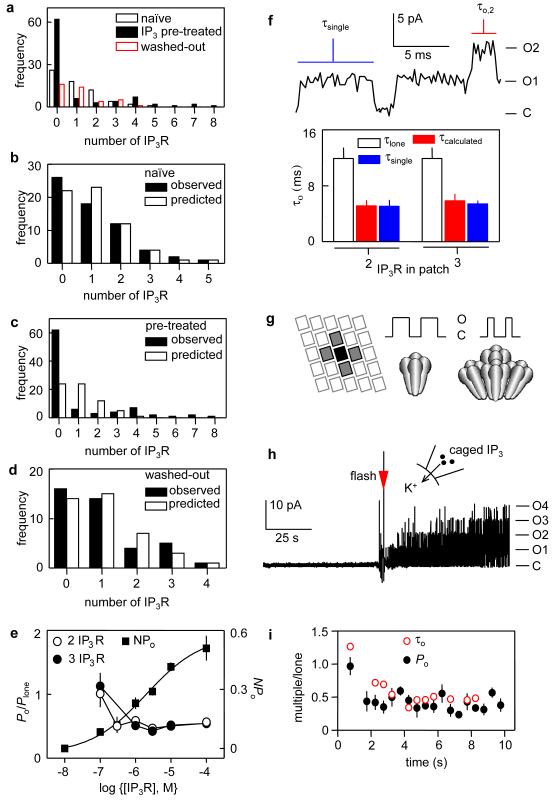
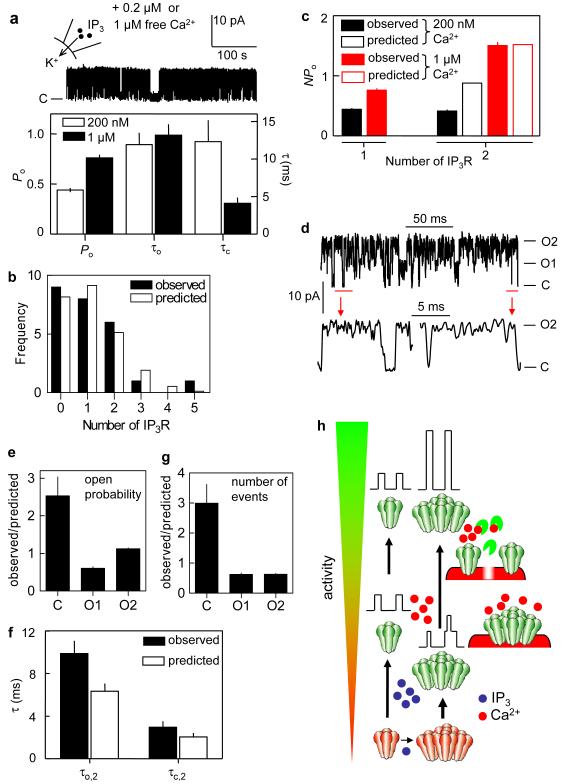
The versatility of Ca2+ signals derives from their spatio-temporal organization. For Ca2+ signals initiated by inositol-1,4,5-trisphosphate (InsP3), this requires local interactions between InsP3 receptors (InsP3Rs) mediated by their rapid stimulation and slower inhibition\ by cytosolic Ca2+. This allows hierarchical recruitment of Ca2+ release events as the InsP3 concentration increases. Single InsP3Rs respond first, then clustered InsP3Rs open together giving a local 'Ca2+ puff', and as puffs become more frequent they ignite regenerative Ca2+ waves. Using nuclear patch-clamp recording, here we demonstrate that InsP3Rs are initially randomly distributed with an estimated separation of 1 m. Low concentrations of InsP3 cause InsP3Rs to aggregate rapidly and reversibly into small clusters of about four closely associated InsP3Rs. At resting cytosolic [Ca2+], clustered InsP3Rs open independently, but with lower open probability, shorter open time, and less InsP3 sensitivity than lone InsP3Rs. Increasing cytosolic [Ca2+] reverses the inhibition caused by clustering, InsP3R gating becomes coupled, and the duration of multiple openings is prolonged. Clustering both exposes InsP3Rs to local Ca2+ rises and increases the effects of Ca2+. Dynamic regulation of clustering by InsP3 retunes InsP3R sensitivity to InsP3 and Ca2+, facilitating hierarchical recruitment of the elementary events that underlie all InsP3-evoked Ca2+ signals.
Figures
Comment in
-
InsP3R channel gating altered by clustering?Nature. 2011 Oct 12;478(7368):E1-2; discussion E2-3. doi: 10.1038/nature10493. Nature. 2011. PMID: 21993761 Free PMC article. No abstract available.
References
-
- Berridge MJ, Lipp P, Bootman MD. The versatility and universality of calcium signalling. Nature Rev. Mol. Cell Biol. 2000;1:11–21. - PubMed
-
- Rizzuto R, Pozzan T. Microdomains of intracellular Ca2+: molecular determinants and functional consequences. Physiol. Rev. 2006;86:369–408. - PubMed
-
- Bootman MD, Berridge MJ, Lipp P. Cooking with calcium: the recipes for composing global signals from elementary events. Cell. 1997;91:367–373. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
