

Early iron-deficiency-induced transcriptional changes in Arabidopsis roots as revealed by microarray analyses
- PMID: 19348669
- PMCID: PMC2676303
- DOI: 10.1186/1471-2164-10-147
Early iron-deficiency-induced transcriptional changes in Arabidopsis roots as revealed by microarray analyses
Abstract
Background: Iron (Fe) is an essential nutrient in plants and animals, and Fe deficiency results in decreased vitality and performance. Due to limited bio-availability of Fe, plants have evolved sophisticated adaptive alterations in development, biochemistry and metabolism that are mainly regulated at the transcriptional level. We have investigated the early transcriptional response to Fe deficiency in roots of the model plant Arabidopsis, using a hydroponic system that permitted removal of Fe from the nutrient solution within seconds and transferring large numbers of plants with little or no mechanical damage to the root systems. We feel that this experimental approach offers significant advantages over previous and recent DNA microarray investigations of the Fe-deficiency response by increasing the resolution of the temporal response and by decreasing non-Fe deficiency-induced transcriptional changes, which are common in microarray analyses.
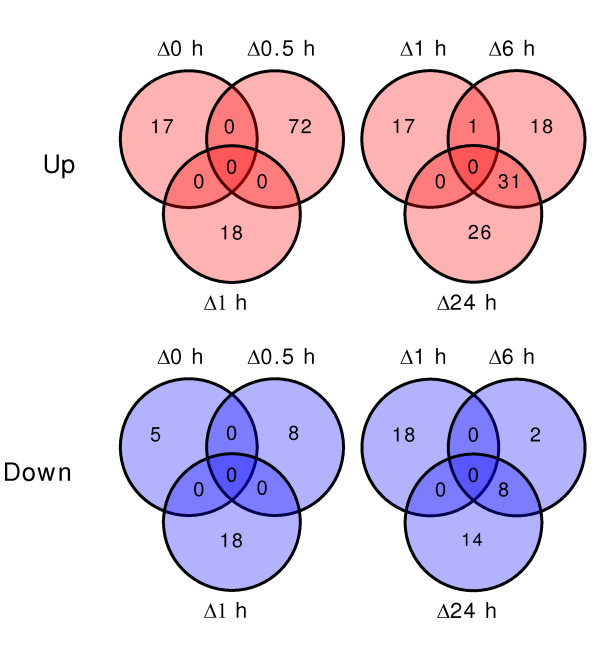
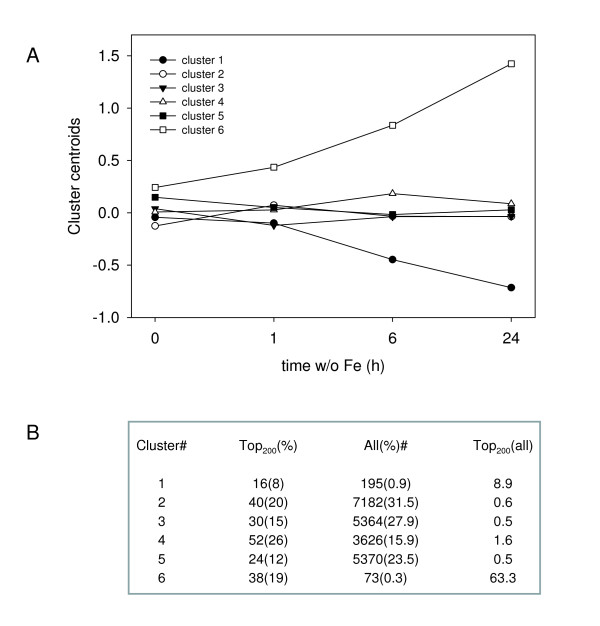
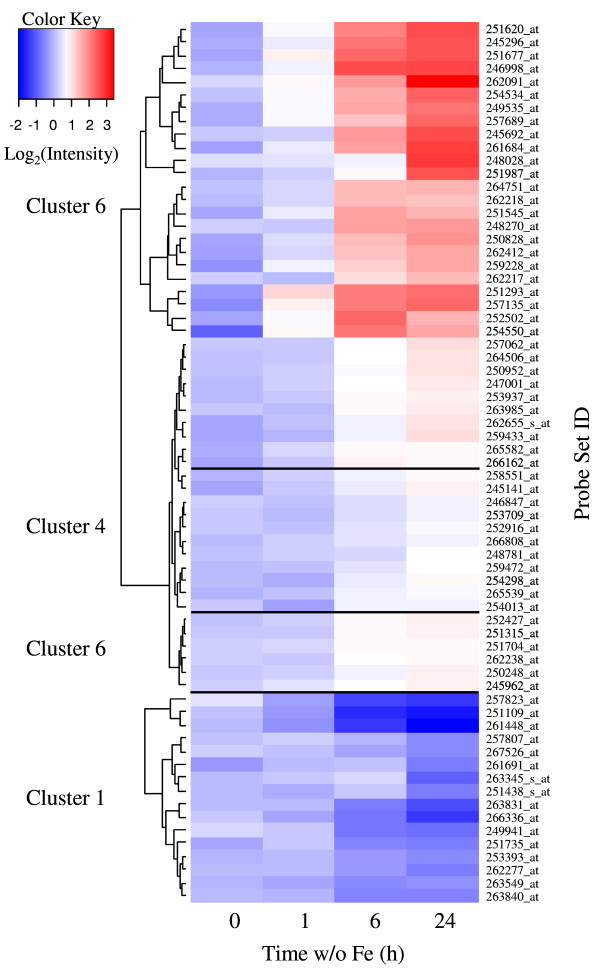
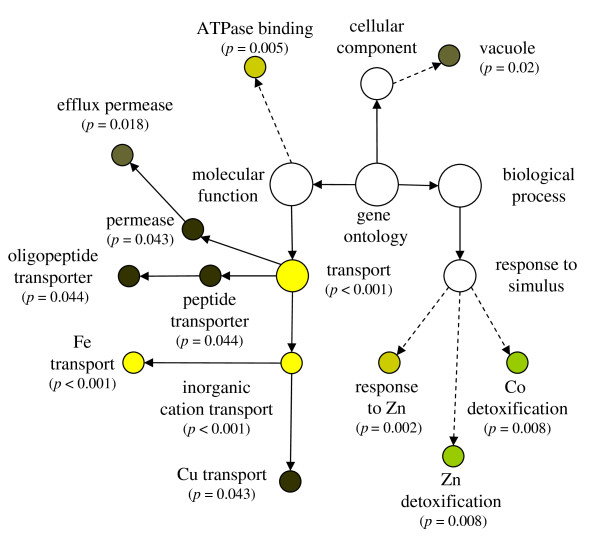
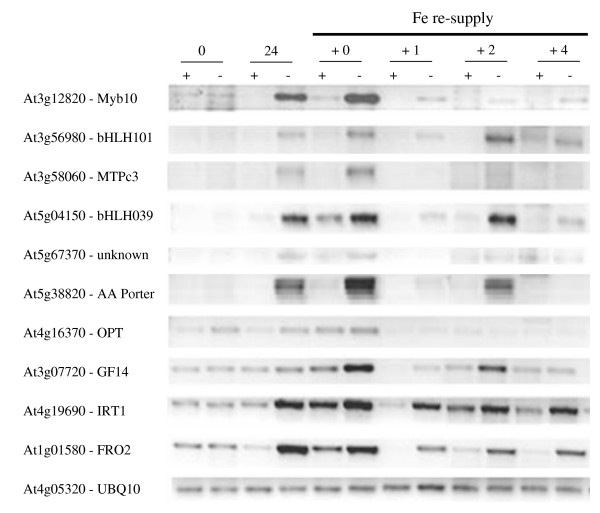
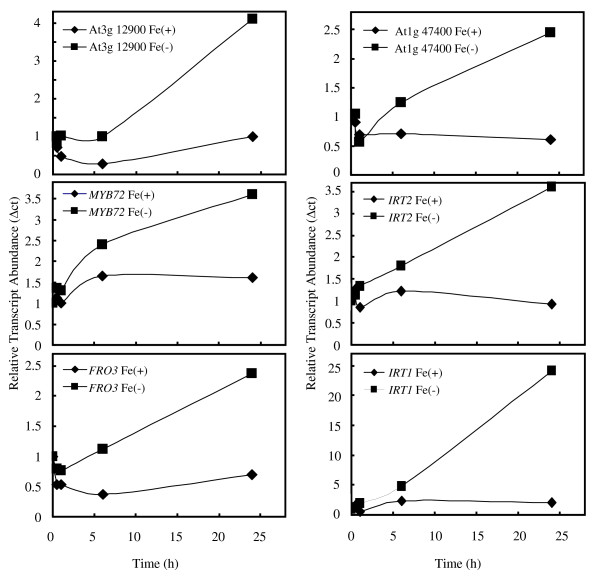
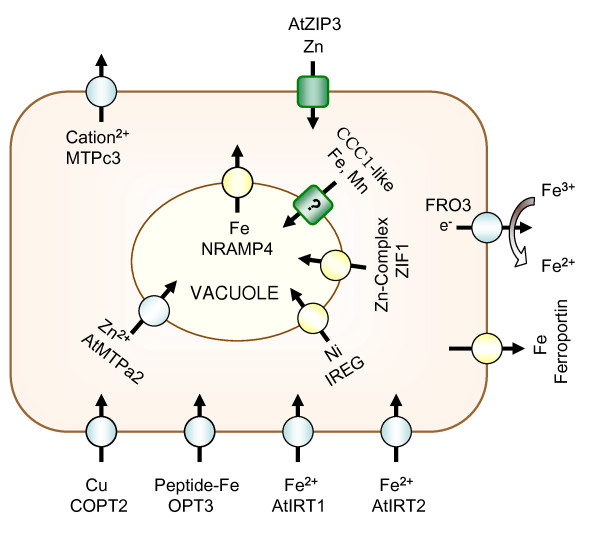
Results: The expression of sixty genes were changed after 6 h of Fe deficiency and 65% of these were found to overlap with a group of seventy-nine genes that were altered after 24 h. A disproportionally high number of transcripts encoding ion transport proteins were found, which function to increase the Fe concentration and decrease the zinc (Zn) concentration in the cytosol. Analysis of global changes in gene expression revealed that changes in Fe availability were associated with the differential expression of genes that encode transporters with presumed function in uptake and distribution of transition metals other than Fe. It appeared that under conditions of Fe deficiency, the capacity for Zn uptake increased, most probably the result of low specificity of the Fe transporter IRT1 that was induced upon Fe deficiency. The transcriptional regulation of several Zn transports under Fe deficiency led presumably to the homeostatic regulation of the cytosolic concentration of Zn and of other transition metal ions such as Mn to avoid toxicity.
Conclusion: The genomic information obtained from this study gives insights into the rapid transcriptional responses to Fe shortage in plants, and is important for understanding how changes in nutrient availability are translated into responses that help to avoid imbalances in ion distribution. We further identified rapidly induced or repressed genes with potential roles in perception and signaling during Fe deficiency which may aid in the elucidation of these processes.
Figures

References
-
- Marschner H. Mineral Nutrition of Higher Plants. 2. London: Academic Press Ltd; 1995.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
