

Macroscopic models of local field potentials and the apparent 1/f noise in brain activity
- PMID: 19348744
- PMCID: PMC2711281
- DOI: 10.1016/j.bpj.2008.12.3951
Macroscopic models of local field potentials and the apparent 1/f noise in brain activity
Abstract
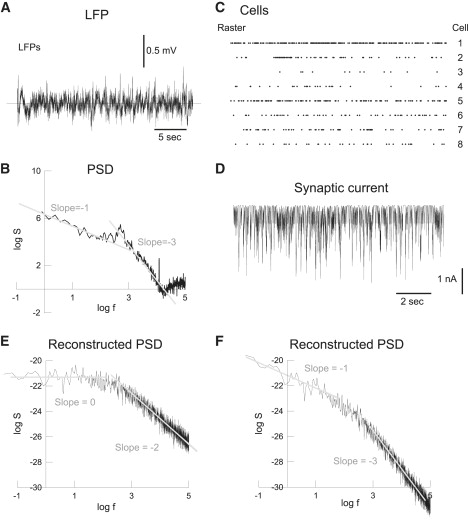
The power spectrum of local field potentials (LFPs) has been reported to scale as the inverse of the frequency, but the origin of this 1/f noise is at present unclear. Macroscopic measurements in cortical tissue demonstrated that electric conductivity (as well as permittivity) is frequency-dependent, while other measurements failed to evidence any dependence on frequency. In this article, we propose a model of the genesis of LFPs that accounts for the above data and contradictions. Starting from first principles (Maxwell equations), we introduce a macroscopic formalism in which macroscopic measurements are naturally incorporated, and also examine different physical causes for the frequency dependence. We suggest that ionic diffusion primes over electric field effects, and is responsible for the frequency dependence. This explains the contradictory observations, and also reproduces the 1/f power spectral structure of LFPs, as well as more complex frequency scaling. Finally, we suggest a measurement method to reveal the frequency dependence of current propagation in biological tissue, and which could be used to directly test the predictions of this formalism.
Figures




References
-
- Bédard C., Kröger H., Destexhe A. Does the 1/f frequency scaling of brain signals reflect self-organized critical states? Phys. Rev. Lett. 2006;97:118102. - PubMed
-
- Novikov E., Novikov A., Shannahoff-Khalsa D., Schwartz B., Wright J. Scale-similar activity in the brain. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Topics. 1997;56:R2387–R2389.
-
- Pritchard W.S. The brain in fractal time: 1/f-like power spectrum scaling of the human electroencephalogram. Int. J. Neurosci. 1992;66:119–129. - PubMed
-
- Jensen H.J. Cambridge University Press; Cambridge, UK: 1998. Self-Organized Criticality: Emergent Complex Behavior in Physical and Biological Systems.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
