

Mechanisms of receptor/coreceptor-mediated entry of enveloped viruses
- PMID: 19348746
- PMCID: PMC2711287
- DOI: 10.1016/j.bpj.2009.01.018
Mechanisms of receptor/coreceptor-mediated entry of enveloped viruses
Abstract
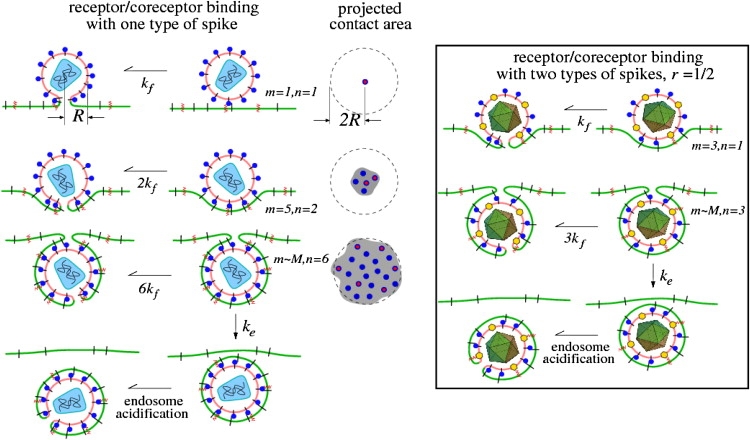
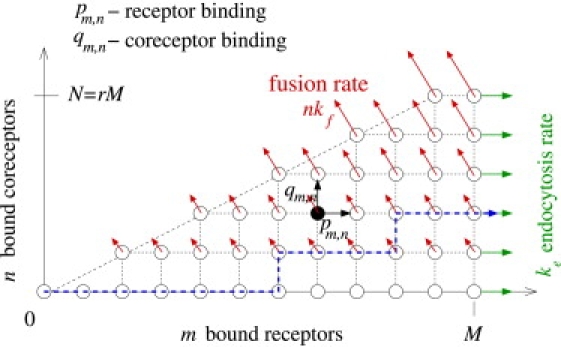
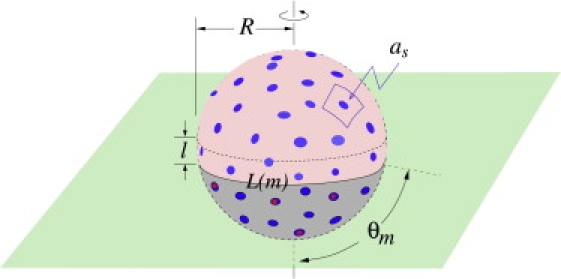
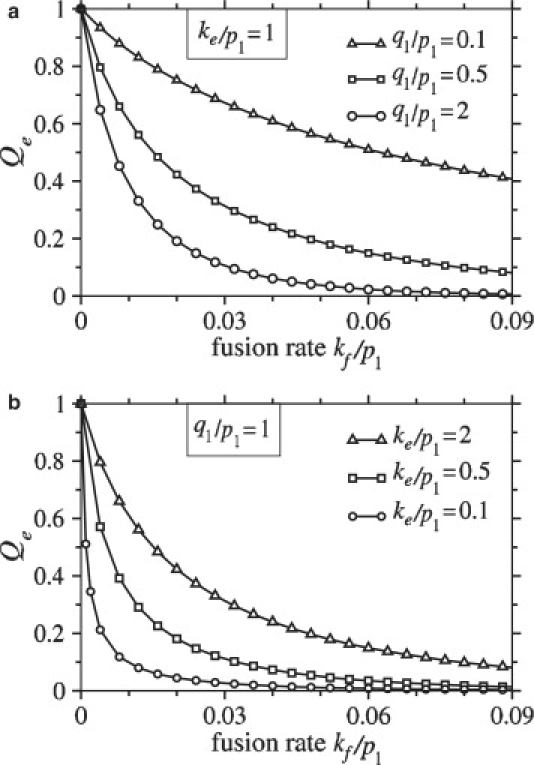
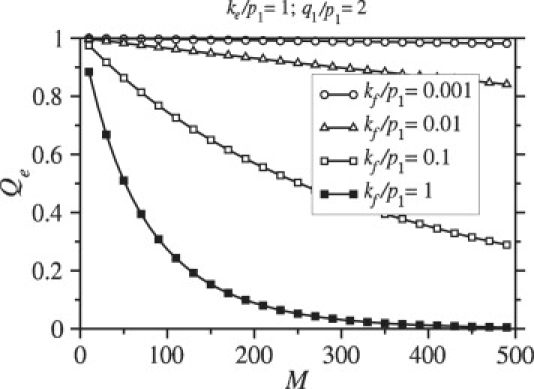
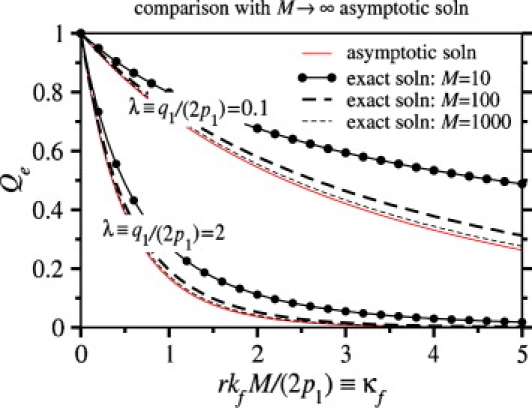
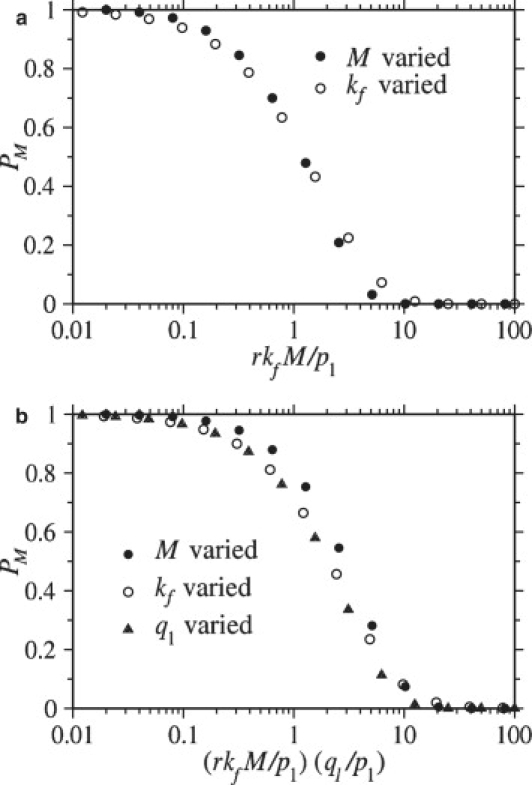
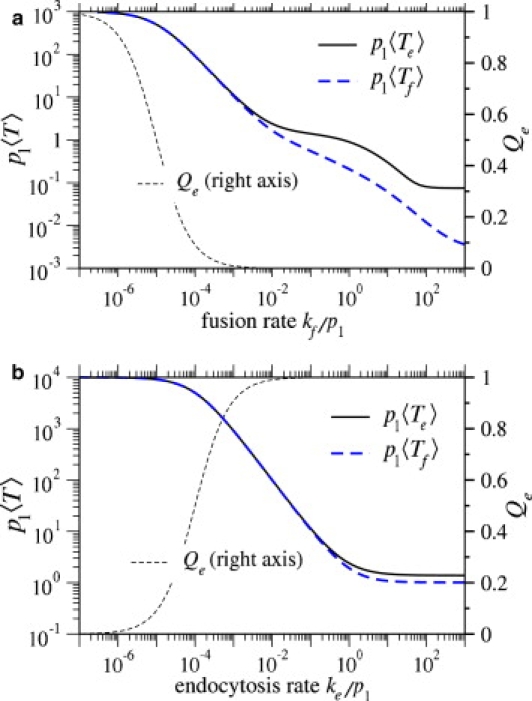
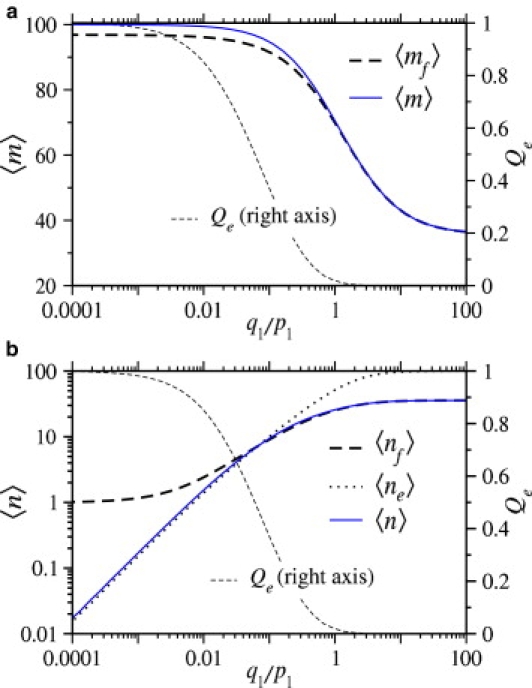
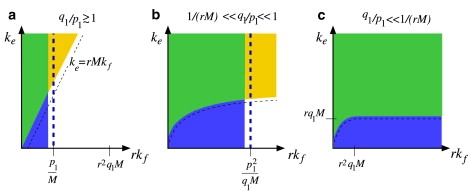
Enveloped viruses enter host cells either through endocytosis, or by direct fusion of the viral envelope and the membrane of the host cell. However, some viruses, such as HIV-1, HSV-1, and Epstein-Barr can enter a cell through either mechanism, with the choice of pathway often a function of the ambient physical chemical conditions, such as temperature and pH. We develop a stochastic model that describes the entry process at the level of binding of viral glycoprotein spikes to cell membrane receptors and coreceptors. In our model, receptors attach the cell membrane to the viral membrane, while subsequent binding of coreceptors enables fusion. The model quantifies the competition between fusion and endocytotic entry pathways. Relative probabilities for each pathway are computed numerically, as well as analytically in the high viral spike density limit. We delineate parameter regimes in which fusion or endocytosis is dominant. These parameters are related to measurable and potentially controllable quantities such as membrane bending rigidity and receptor, coreceptor, and viral spike densities. Experimental implications of our mechanistic hypotheses are proposed and discussed.
Figures
Similar articles
-
Stochastic entry of enveloped viruses: fusion versus endocytosis.Biophys J. 2007 Aug 15;93(4):1116-23. doi: 10.1529/biophysj.107.106708. Epub 2007 May 18. Biophys J. 2007. PMID: 17513379 Free PMC article.
-
Cell entry of enveloped viruses.Adv Genet. 2011;73:121-83. doi: 10.1016/B978-0-12-380860-8.00004-5. Adv Genet. 2011. PMID: 21310296 Free PMC article. Review.
-
Mechanisms of coronavirus cell entry mediated by the viral spike protein.Viruses. 2012 Jun;4(6):1011-33. doi: 10.3390/v4061011. Epub 2012 Jun 20. Viruses. 2012. PMID: 22816037 Free PMC article. Review.
-
Two Sides to Every Story: Herpes Simplex Type-1 Viral Glycoproteins gB, gD, gH/gL, gK, and Cellular Receptors Function as Key Players in Membrane Fusion.Viruses. 2021 Sep 16;13(9):1849. doi: 10.3390/v13091849. Viruses. 2021. PMID: 34578430 Free PMC article. Review.
-
Diffusion-dependent mechanisms of receptor engagement and viral entry.J Phys Chem B. 2010 Nov 25;114(46):15403-12. doi: 10.1021/jp1080725. Epub 2010 Nov 1. J Phys Chem B. 2010. PMID: 21038861
Cited by
-
Stochastic model-assisted development of efficient low-dose viral transduction in microfluidics.Biophys J. 2013 Feb 19;104(4):934-42. doi: 10.1016/j.bpj.2012.12.049. Biophys J. 2013. PMID: 23442972 Free PMC article.
-
Broadly Neutralizing Antibodies against HIV-1 As a Novel Aspect of the Immune Response.Acta Naturae. 2015 Oct-Dec;7(4):11-21. Acta Naturae. 2015. PMID: 26798488 Free PMC article.
-
How Viruses Use the VCP/p97 ATPase Molecular Machine.Viruses. 2021 Sep 21;13(9):1881. doi: 10.3390/v13091881. Viruses. 2021. PMID: 34578461 Free PMC article. Review.
-
Antiviral Protein-Protein Interaction Inhibitors.J Med Chem. 2024 Mar 14;67(5):3205-3231. doi: 10.1021/acs.jmedchem.3c01543. Epub 2024 Feb 23. J Med Chem. 2024. PMID: 38394369 Free PMC article. Review.
-
Lipid rafts as viral entry routes and immune platforms: A double-edged sword in SARS-CoV-2 infection?Biochim Biophys Acta Mol Cell Biol Lipids. 2022 Jun;1867(6):159140. doi: 10.1016/j.bbalip.2022.159140. Epub 2022 Mar 4. Biochim Biophys Acta Mol Cell Biol Lipids. 2022. PMID: 35248801 Free PMC article. Review.
References
-
- Duzgunes N., Pedroso de Lima M.C., Stamatatos L., Flasher D., Alford D. Fusion activity and inactivation of influenza virus: kinetics of low pH-induced fusion with cultured cells. J. Gen. Virol. 1992;73:27–37. - PubMed
-
- Marsh M., Bron R. SFV infection in CHO cells: cell-type specific restrictions to productive virus entry at the cell surface. J. Cell Sci. 1997;110:95–103. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
