

Electrically silent divalent cation entries in resting and active voltage-controlled muscle fibers
- PMID: 19348748
- PMCID: PMC2711298
- DOI: 10.1016/j.bpj.2009.01.008
Electrically silent divalent cation entries in resting and active voltage-controlled muscle fibers
Abstract
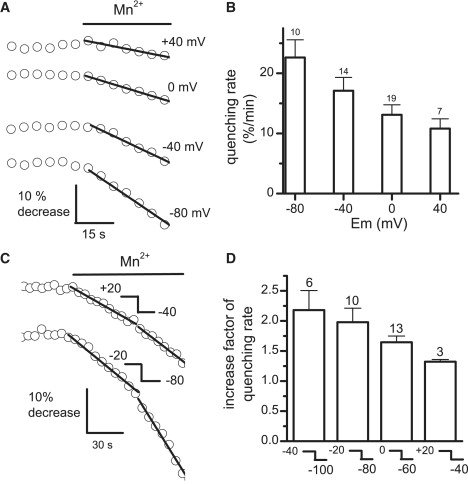
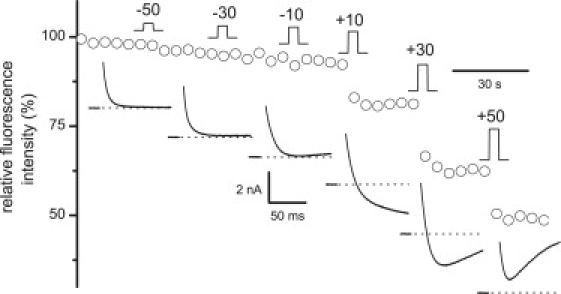
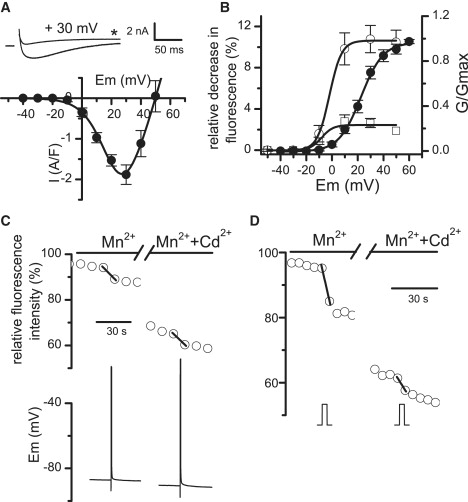
Ca2+ is known to enter skeletal muscle at rest and during activity. Except for the well-characterized Ca2+ entry through L-type channels, pathways involved in these Ca2+ entries remain elusive in adult muscle. This study investigates Ca2+ influx at rest and during activity using the method of Mn2+ quenching of fura-2 fluorescence on voltage-controlled adult skeletal muscle cells. Resting rate of Mn2+ influx depended on external [Mn2+] and membrane potential. At -80 mV, replacement of Mg2+ by Mn2+ gave rise to an outward current associated with an increase in cell input resistance. Calibration of fura-2 response indicated that Mn2+ influx was too small to be resolved as a macroscopic current. Partial depletion of the sarcoplasmic reticulum induced by a train of action potentials in the presence of cyclopiazonic acid led to a slight increase in resting Mn2+ influx but no change in cell input resistance and membrane potential. Trains of action potentials considerably increased Mn2+ entry through an electrically silent pathway independent of L-type channels, which provided 24% of the global Mn2+ influx at +30 mV under voltage-clamp conditions. Within this context, the nature and the physiological role of the Ca2+ pathways involved during muscle excitation still remain open questions.
Figures




References
-
- Gailly P. New aspects of calcium signaling in skeletal muscle cells: implications in Duchenne muscular dystrophy. Biochim. Biophys. Acta. 2002;1600:38–44. - PubMed
-
- Haws C.M., Lansman J.B. Developmental regulation of mechanosensitive calcium channels in skeletal muscle from normal and mdx mice. Proc. Biol. Sci. 1991;245:173–177. - PubMed
-
- Hopf F.W., Turner P.R., Denetclaw W.F., Jr., Reddy P., Steinhardt R.A. A critical evaluation of resting intracellular free calcium regulation in dystrophic mdx muscle. Am. J. Physiol. 1996;271:C1325–C1339. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
