

Calmodulin mediates the Ca2+-dependent regulation of Cx44 gap junctions
- PMID: 19348766
- PMCID: PMC2711270
- DOI: 10.1016/j.bpj.2008.12.3941
Calmodulin mediates the Ca2+-dependent regulation of Cx44 gap junctions
Abstract
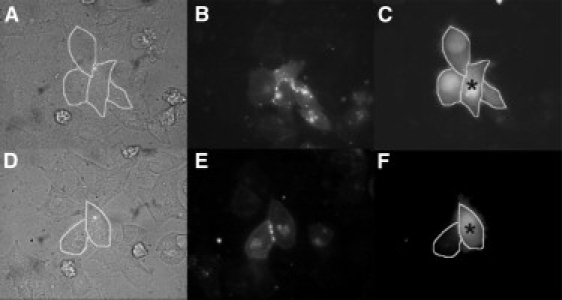
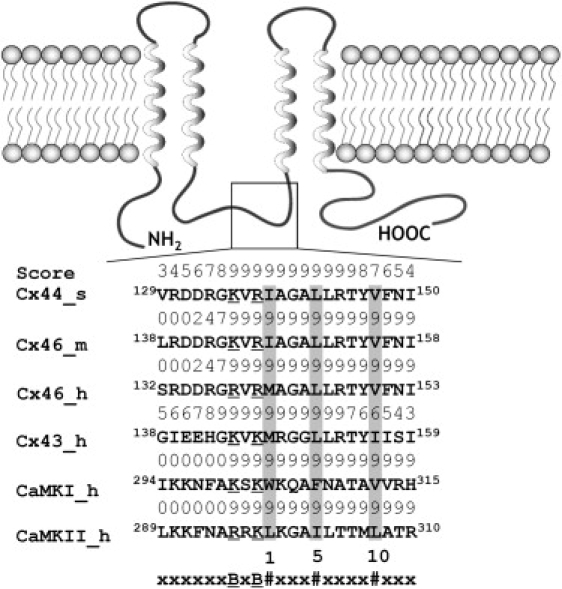
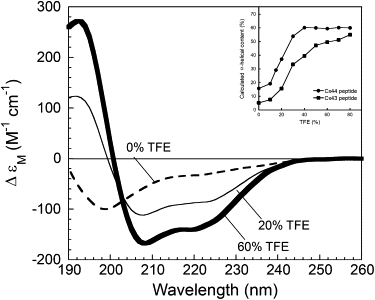
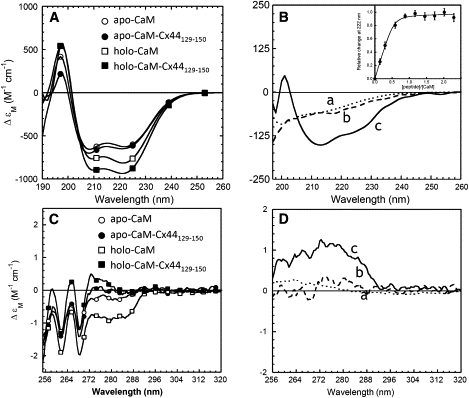
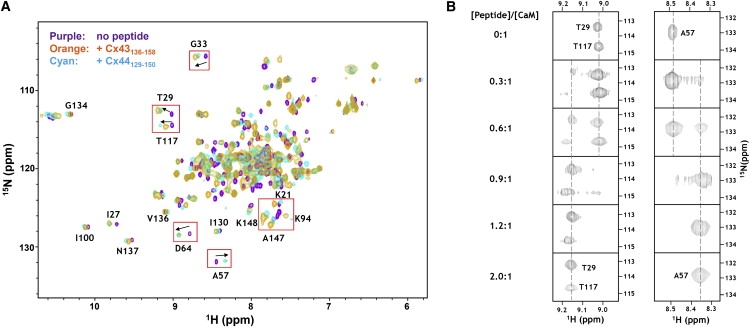
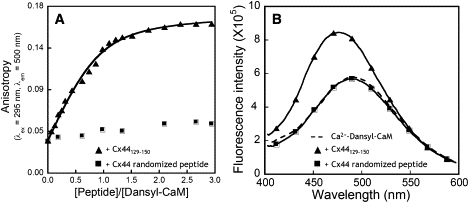
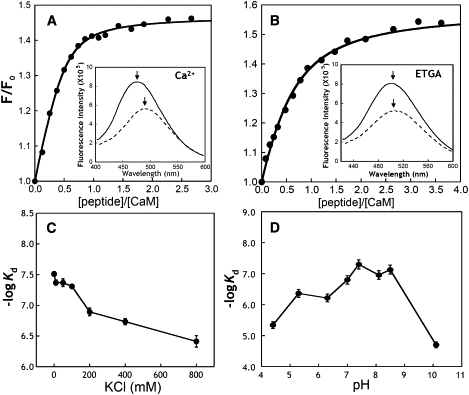
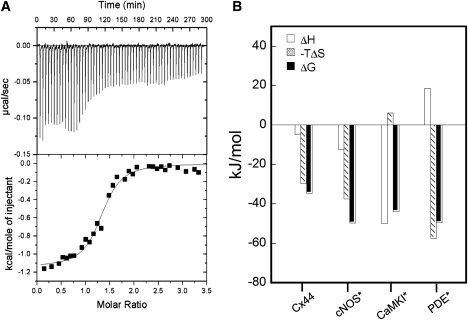
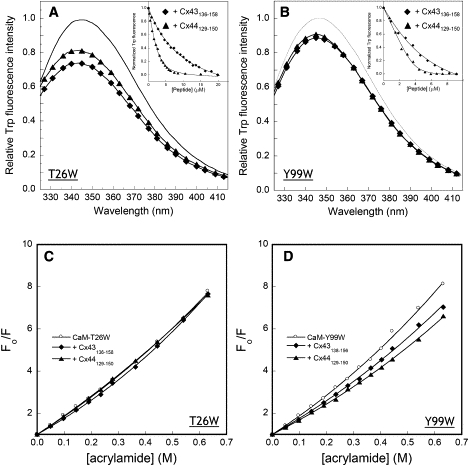
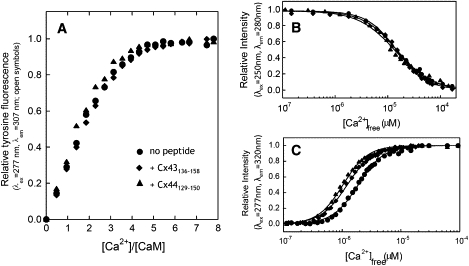
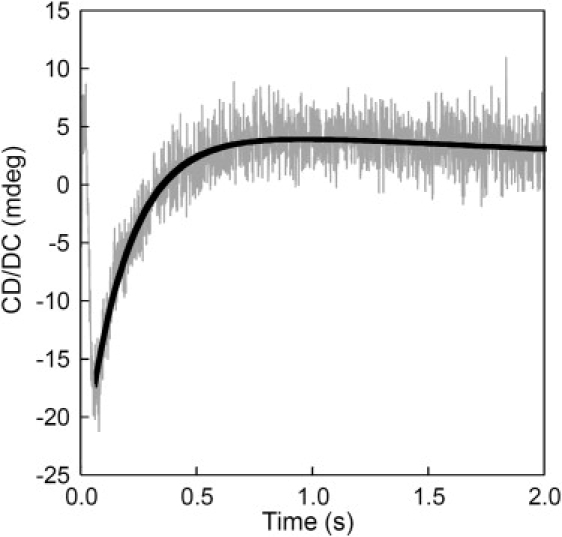
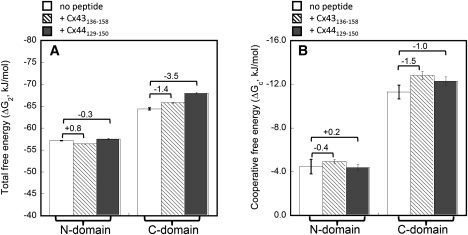
We have shown previously that the Ca2+-dependent inhibition of lens epithelial cell-to-cell communication is mediated in part by the direct association of calmodulin (CaM) with connexin43 (Cx43), the major connexin in these cells. We now show that elevation of [Ca2+](i) in HeLa cells transfected with the lens fiber cell gap junction protein sheep Cx44 also results in the inhibition of cell-to-cell dye transfer. A peptide comprising the putative CaM binding domain (aa 129-150) of the intracellular loop region of this connexin exhibited a high affinity, stoichiometric interaction with Ca2+-CaM. NMR studies indicate that the binding of Cx44 peptide to CaM reflects a classical embracing mode of interaction. The interaction is an exothermic event that is both enthalpically and entropically driven in which electrostatic interactions play an important role. The binding of the Cx44 peptide to CaM increases the CaM intradomain cooperativity and enhances the Ca2+-binding affinities of the C-domain of CaM more than twofold by slowing the rate of Ca2+ release from the complex. Our data suggest a common mechanism by which the Ca2+-dependent inhibition of the alpha-class of gap junction proteins is mediated by the direct association of an intracellular loop region of these proteins with Ca2+-CaM.
Figures
References
-
- Mathias R.T., Rae J.L., Baldo G.J. Physiological properties of the normal lens. Physiol. Rev. 1997;77:21–50. - PubMed
-
- Eiberger J., Degen J., Romualdi A., Deutsch U., Willecke K. Connexin genes in the mouse and human genome. Cell Commun. Adhes. 2001;8:163–165. - PubMed
-
- Eastman S.D., Chen T.H., Falk M.M., Mendelson T.C., Iovine M.K. Phylogenetic analysis of three complete gap junction gene families reveals lineage-specific duplications and highly supported gene classes. Genomics. 2006;87:265–274. - PubMed
-
- Krutovskikh V., Yamasaki H. Connexin gene mutations in human genetic diseases. Mutat. Res. 2000;462:197–207. - PubMed
-
- Gao Y., Spray D.C. Structural changes in lenses of mice lacking the gap junction protein connexin43. Invest. Ophthalmol. Vis. Sci. 1998;39:1198–1209. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
