

TRIM32 is an E3 ubiquitin ligase for dysbindin
- PMID: 19349376
- PMCID: PMC2694686
- DOI: 10.1093/hmg/ddp167
TRIM32 is an E3 ubiquitin ligase for dysbindin
Abstract
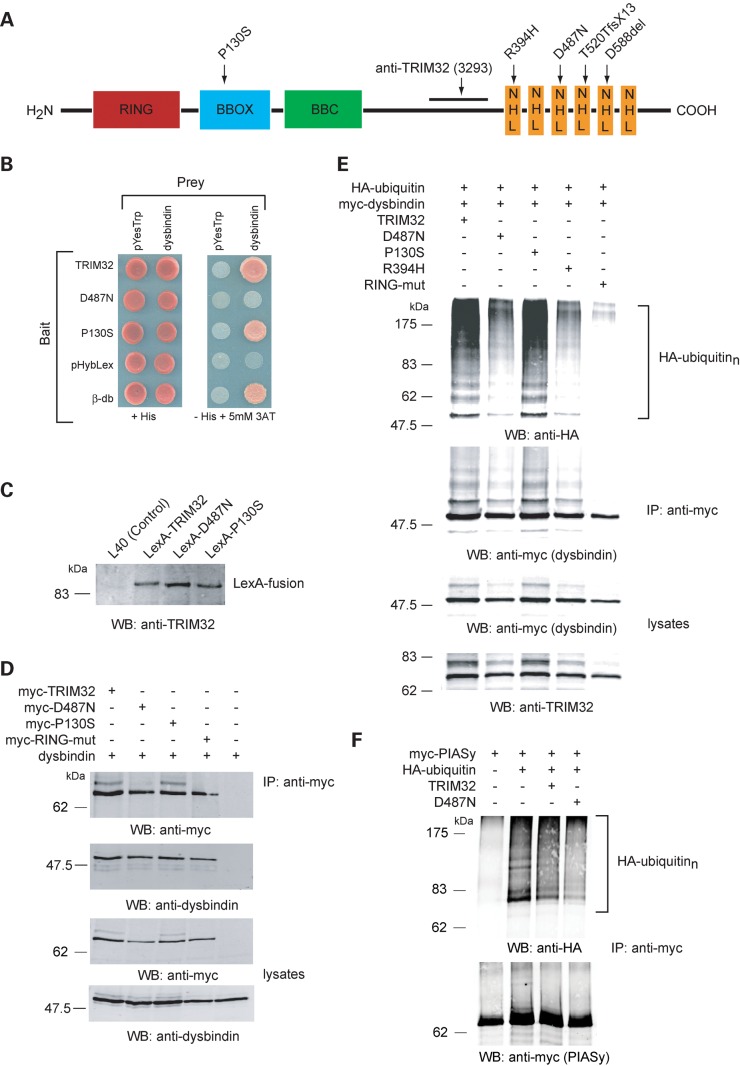
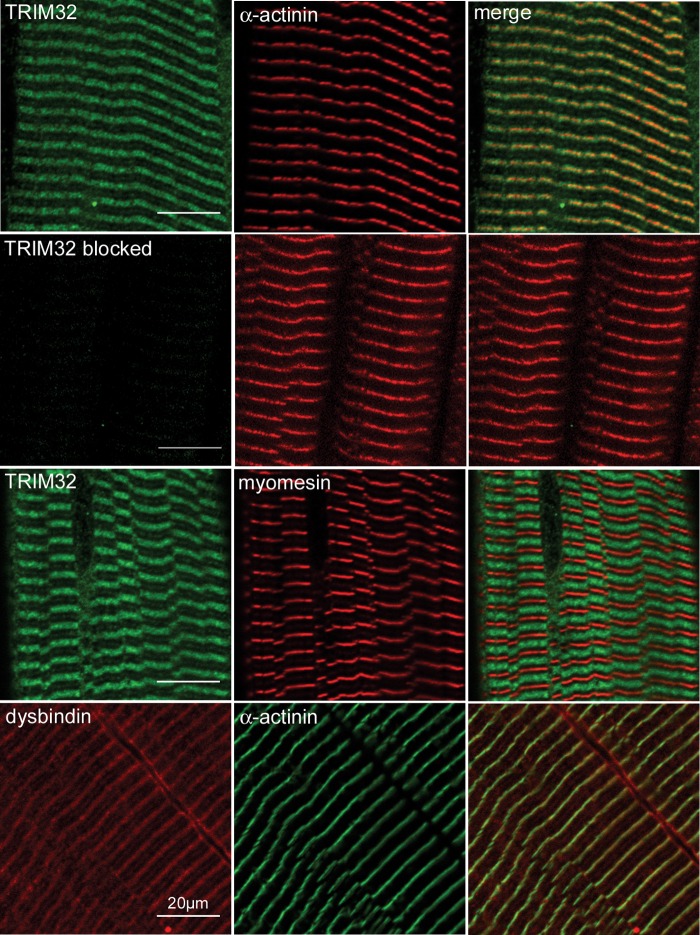
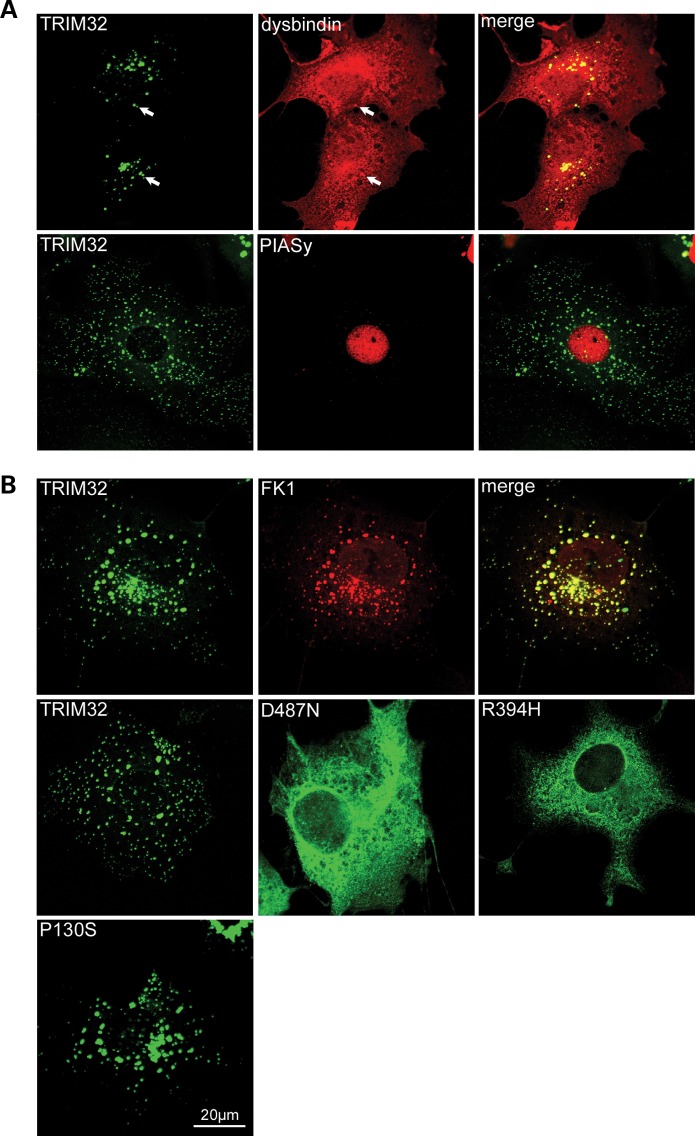
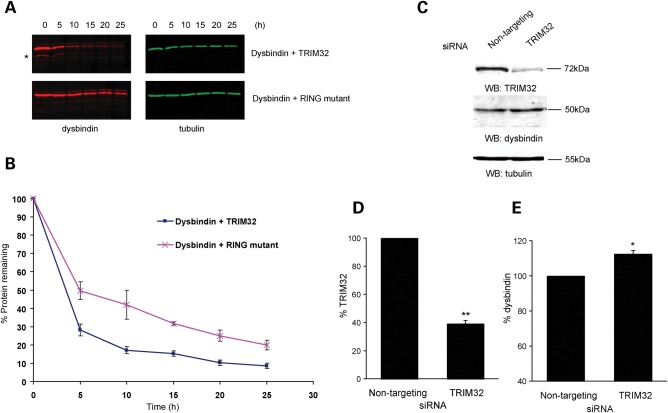
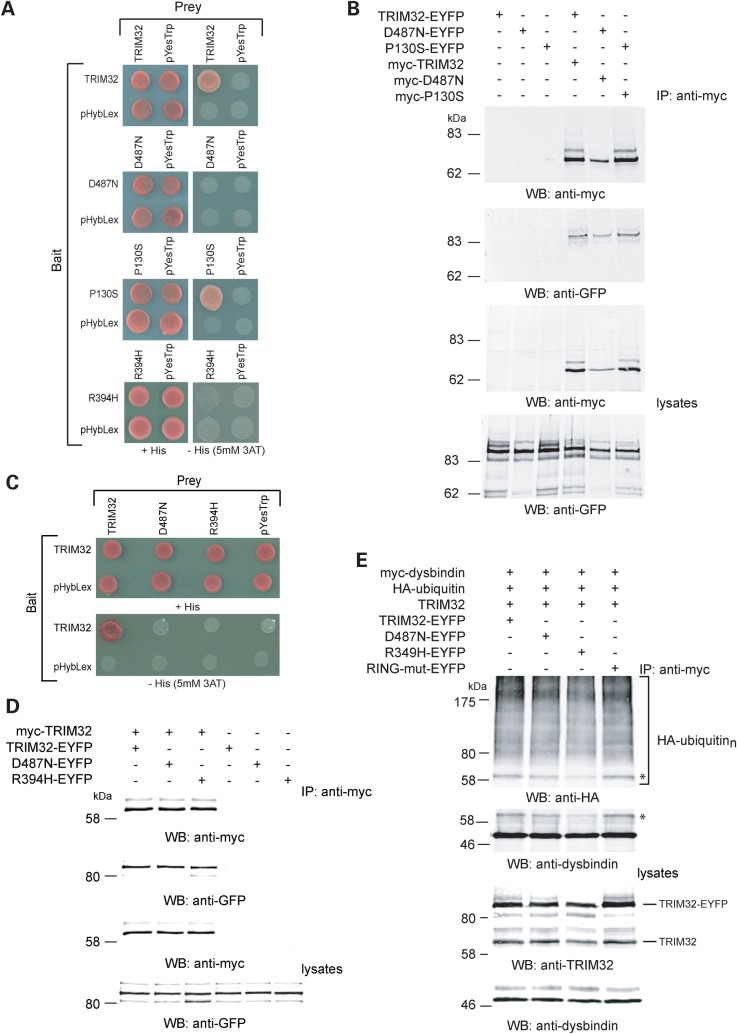
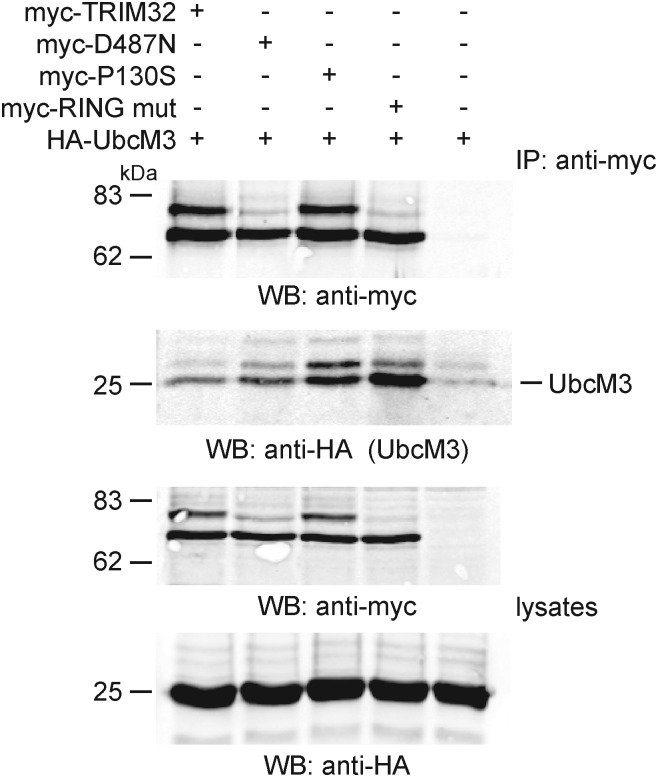
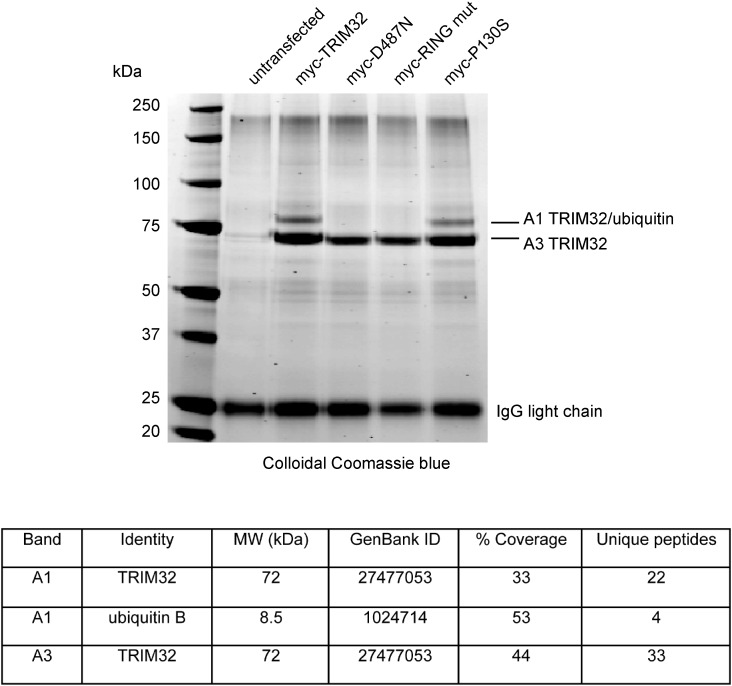
Mutations in the gene encoding tripartite motif protein 32 (TRIM32) cause two seemingly diverse diseases: limb-girdle muscular dystrophy type 2H (LGMD2H) or sarcotubular myopathy (STM) and Bardet-Biedl syndrome type 11(BBS11). Although TRIM32 is involved in protein ubiquitination, its substrates and the molecular consequences of disease-causing mutations are poorly understood. In this paper, we show that TRIM32 is a widely expressed ubiquitin ligase that is localized to the Z-line in skeletal muscle. Using the yeast two-hybrid system, we found that TRIM32 binds and ubiquitinates dysbindin, a protein implicated in the genetic aetiology of schizophrenia, augmenting its degradation. Small-interfering RNA-mediated knock-down of TRIM32 in myoblasts resulted in elevated levels of dysbindin. Importantly, the LGMD2H/STM-associated TRIM32 mutations, D487N and R394H impair ubiquitin ligase activity towards dysbindin and were mislocalized in heterologous cells. These mutants were able to self-associate and also co-immunoprecipitated with wild-type TRIM32 in transfected cells. Furthermore, the D487N mutant could bind to both dysbindin and its E2 enzyme but was defective in monoubiquitination. In contrast, the BBS11 mutant P130S did not show any biochemical differences compared with the wild-type protein. Our data identify TRIM32 as a regulator of dysbindin and demonstrate that the LGMD2H/STM mutations may impair substrate ubiquitination.
Figures
References
-
- Hershko A., Ciechanover A. The ubiquitin system. Annu. Rev. Biochem. 1998;67:425–479. - PubMed
-
- Pickart C.M. Mechanisms underlying ubiquitination. Annu. Rev. Biochem. 2001;70:503–533. - PubMed
-
- Nisole S., Stoye J.P., Saib A. TRIM family proteins: retroviral restriction and antiviral defence. Nat. Rev. Microbiol. 2005;3:799–808. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
