

Alternatively activated macrophage-derived RELM-{alpha} is a negative regulator of type 2 inflammation in the lung
- PMID: 19349464
- PMCID: PMC2715126
- DOI: 10.1084/jem.20082048
Alternatively activated macrophage-derived RELM-{alpha} is a negative regulator of type 2 inflammation in the lung
Erratum in
- J Exp Med. 2009 May 11;206(5):1201
Abstract
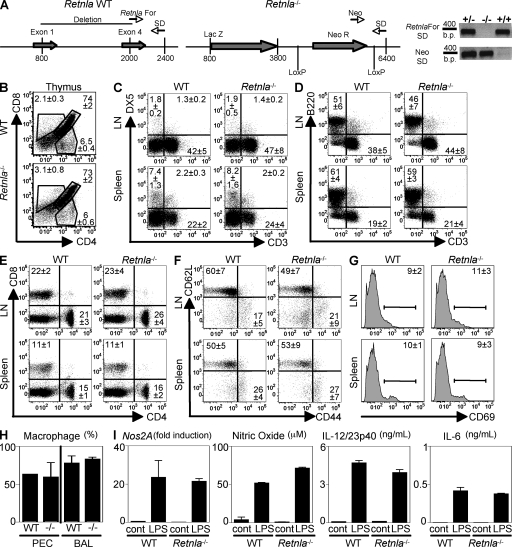
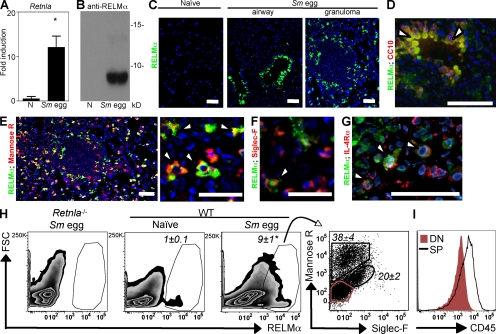
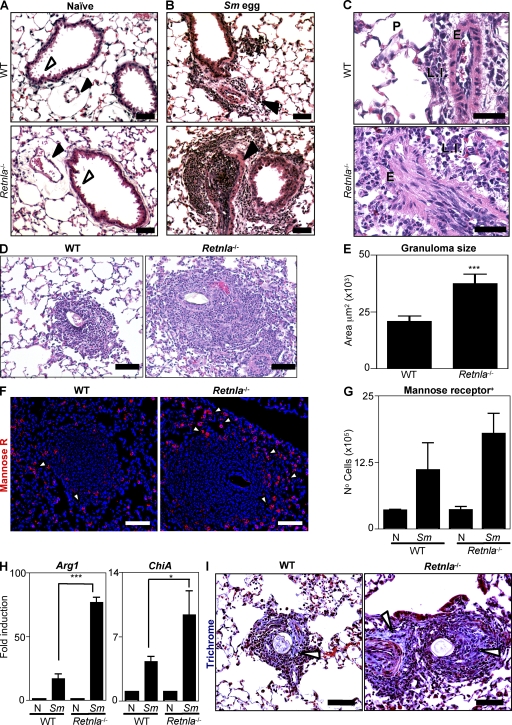
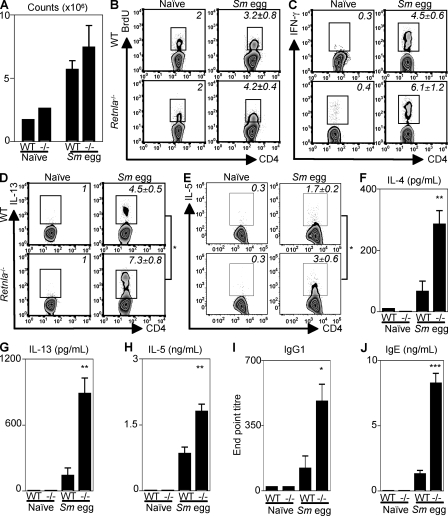
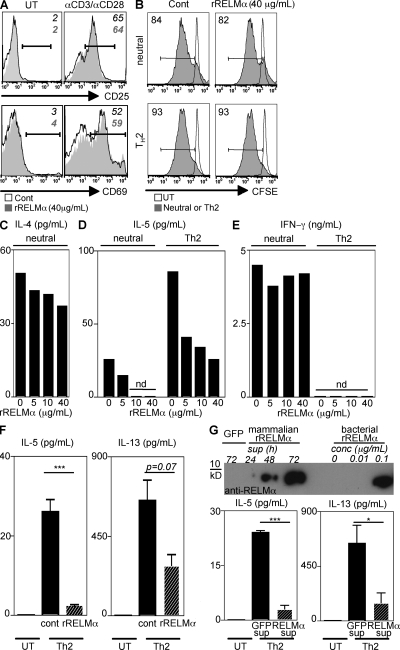
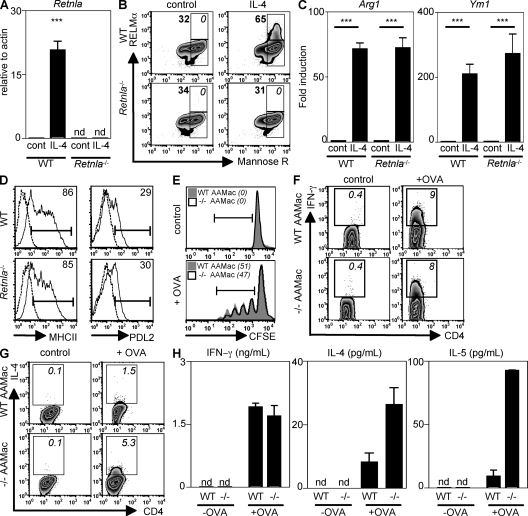
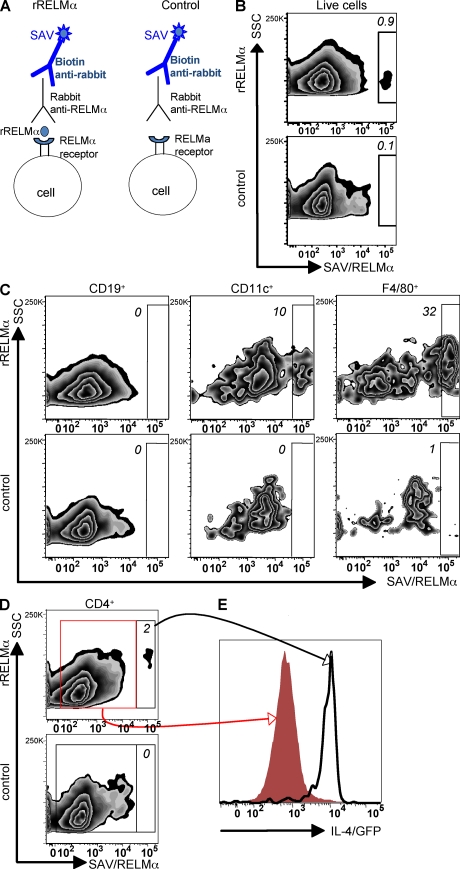
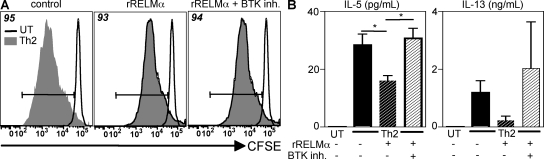
Differentiation and recruitment of alternatively activated macrophages (AAMacs) are hallmarks of several inflammatory conditions associated with infection, allergy, diabetes, and cancer. AAMacs are defined by the expression of Arginase 1, chitinase-like molecules, and resistin-like molecule (RELM) alpha/FIZZ1; however, the influence of these molecules on the development, progression, or resolution of inflammatory diseases is unknown. We describe the generation of RELM-alpha-deficient (Retnla(-/-)) mice and use a model of T helper type 2 (Th2) cytokine-dependent lung inflammation to identify an immunoregulatory role for RELM-alpha. After challenge with Schistosoma mansoni (Sm) eggs, Retnla(-/-) mice developed exacerbated lung inflammation compared with their wild-type counterparts, characterized by excessive pulmonary vascularization, increased size of egg-induced granulomas, and elevated fibrosis. Associated with increased disease severity, Sm egg-challenged Retnla(-/-) mice exhibited elevated expression of pathogen-specific CD4(+) T cell-derived Th2 cytokines. Consistent with immunoregulatory properties, recombinant RELM-alpha could bind to macrophages and effector CD4(+) Th2 cells and inhibited Th2 cytokine production in a Bruton's tyrosine kinase-dependent manner. Additionally, Retnla(-/-) AAMacs promoted exaggerated antigen-specific Th2 cell differentiation. Collectively, these data identify a previously unrecognized role for AAMac-derived RELM-alpha in limiting the pathogenesis of Th2 cytokine-mediated pulmonary inflammation, in part through the regulation of CD4(+) T cell responses.
Figures
References
-
- Janeway C.A., Jr., Medzhitov R. 2002. Innate immune recognition.Annu. Rev. Immunol. 20:197–216 - PubMed
-
- Stuart L.M., Ezekowitz R.A. 2008. Phagocytosis and comparative innate immunity: learning on the fly.Nat. Rev. Immunol. 8:131–141 - PubMed
-
- Lawrence T., Bebien M., Liu G.Y., Nizet V., Karin M. 2005. IKKalpha limits macrophage NF-kappaB activation and contributes to the resolution of inflammation.Nature. 434:1138–1143 - PubMed
-
- Qualls J.E., Kaplan A.M., van Rooijen N., Cohen D.A. 2006. Suppression of experimental colitis by intestinal mononuclear phagocytes.J. Leukoc. Biol. 80:802–815 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI074878/AI/NIAID NIH HHS/United States
- T32AI007532-08/AI/NIAID NIH HHS/United States
- R56 AI032573/AI/NIAID NIH HHS/United States
- DK50306/DK/NIDDK NIH HHS/United States
- AI53825/AI/NIAID NIH HHS/United States
- AI74878/AI/NIAID NIH HHS/United States
- R01 AI061570/AI/NIAID NIH HHS/United States
- AI61570/AI/NIAID NIH HHS/United States
- P30 DK050306/DK/NIDDK NIH HHS/United States
- T32 AI007532/AI/NIAID NIH HHS/United States
- R01 AI053825/AI/NIAID NIH HHS/United States
- R01 AI032573/AI/NIAID NIH HHS/United States
- AI32573/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
