

Drug inhibition of HDAC3 and epigenetic control of differentiation in Apicomplexa parasites
- PMID: 19349466
- PMCID: PMC2715132
- DOI: 10.1084/jem.20082826
Drug inhibition of HDAC3 and epigenetic control of differentiation in Apicomplexa parasites
Abstract
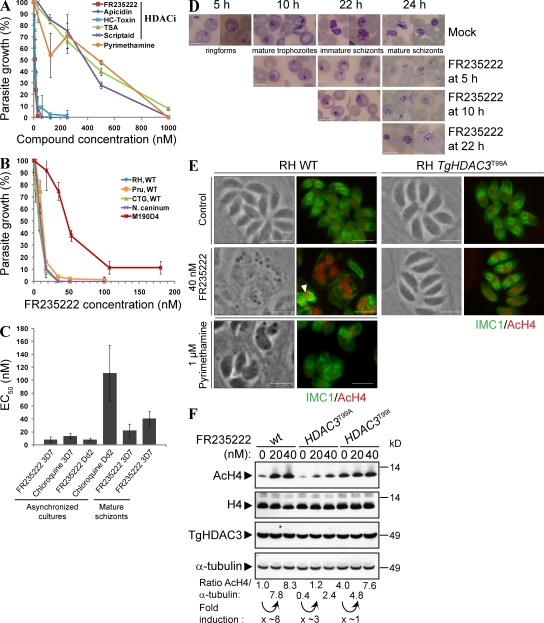
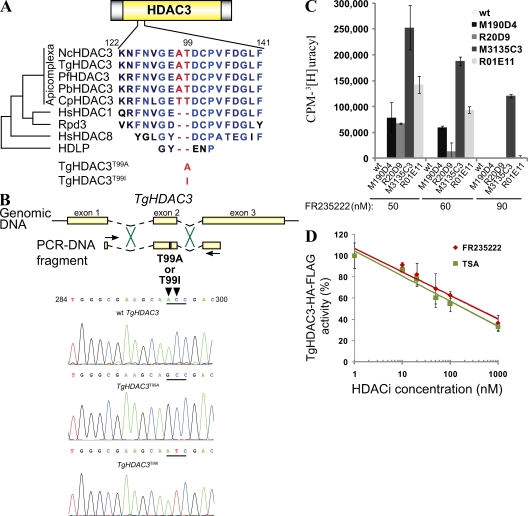
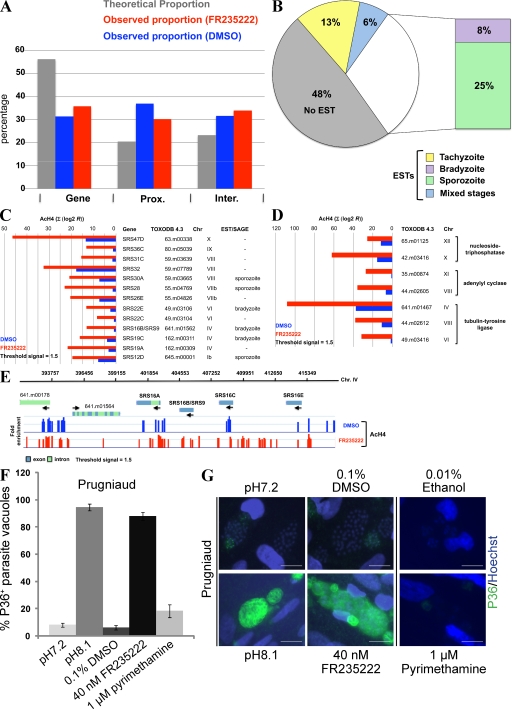
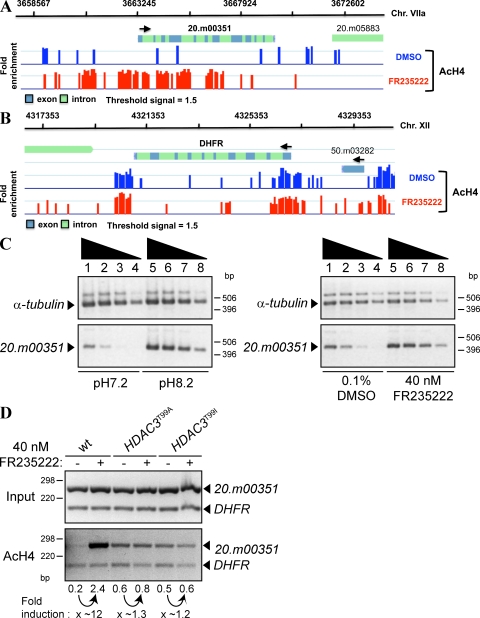
Plasmodium and Toxoplasma are parasites of major medical importance that belong to the Apicomplexa phylum of protozoa. These parasites transform into various stages during their life cycle and express a specific set of proteins at each stage. Although little is yet known of how gene expression is controlled in Apicomplexa, histone modifications, particularly acetylation, are emerging as key regulators of parasite differentiation and stage conversion. We investigated the anti-Apicomplexa effect of FR235222, a histone deacetylase inhibitor (HDACi). We show that FR235222 is active against a variety of Apicomplexa genera, including Plasmodium and Toxoplasma, and is more potent than other HDACi's such as trichostatin A and the clinically relevant compound pyrimethamine. We identify T. gondii HDAC3 (TgHDAC3) as the target of FR235222 in Toxoplasma tachyzoites and demonstrate the crucial role of the conserved and Apicomplexa HDAC-specific residue TgHDAC3 T99 in the inhibitory activity of the drug. We also show that FR235222 induces differentiation of the tachyzoite (replicative) into the bradyzoite (nonreplicative) stage. Additionally, via its anti-TgHDAC3 activity, FR235222 influences the expression of approximately 370 genes, a third of which are stage-specifically expressed. These results identify FR235222 as a potent HDACi of Apicomplexa, and establish HDAC3 as a central regulator of gene expression and stage conversion in Toxoplasma and, likely, other Apicomplexa.
Figures
References
-
- Hakimi M.A., Deitsch K.W. 2007. Epigenetics in Apicomplexa: control of gene expression during cell cycle progression, differentiation and antigenic variation.Curr. Opin. Microbiol. 10:357–362 - PubMed
-
- Maubon D., Ajzenberg D., Brenier-Pinchart M.P., Darde M.L., Pelloux H. 2008. What are the respective host and parasite contributions to toxoplasmosis? Trends Parasitol. 24:299–303 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
