

Inhibitory contribution to suprathreshold corticostriatal responses: an experimental and modeling study
- PMID: 19350384
- PMCID: PMC11505830
- DOI: 10.1007/s10571-009-9394-2
Inhibitory contribution to suprathreshold corticostriatal responses: an experimental and modeling study
Abstract
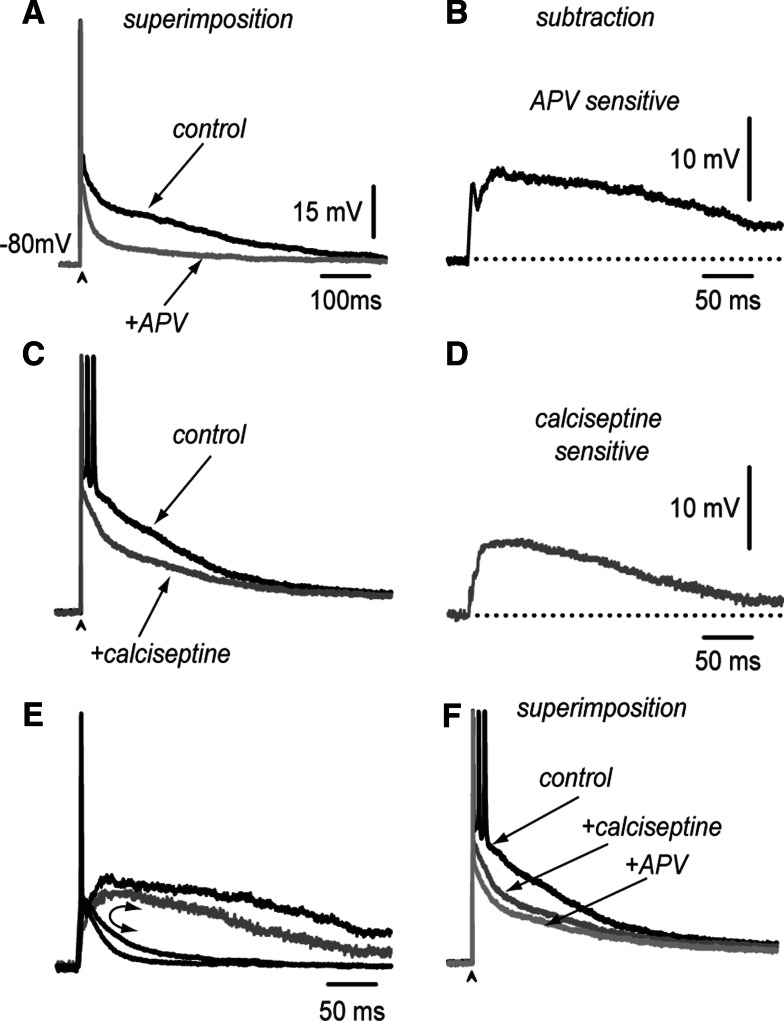
Neostriatal neurons may undergo events of spontaneous synchronization as those observed in recurrent networks of excitatory neurons, even when cortical afferents are transected. It is necessary to explain these events because the neostriatum is a recurrent network of inhibitory neurons. Synchronization of neuronal activity may be caused by plateau-like depolarizations. Plateau-like orthodromic depolarizations that resemble up-states in medium spiny neostriatal neurons (MSNs) may be induced by a single corticostriatal suprathreshold stimulus. Slow synaptic depolarizations may last hundreds of milliseconds, decay slower than the monosynaptic glutamatergic synaptic potentials that induce them, and sustain repetitive firing. Because inhibitory inputs impinging onto MSNs have a reversal potential above the resting membrane potential but below the threshold for firing, they conform a type of "shunting inhibition". This work asks if shunting GABAergic inputs onto MSNs arrive asynchronously enough as to help in sustaining the plateau-like corticostriatal response after a single cortical stimulus. This may help to begin explaining autonomous processing in the striatal micro-circuitry in the presence of a tonic excitatory drive and independently of spatio-temporally organized inputs. It is shown here that besides synaptic currents from AMPA/KA- and NMDA-receptors, as well as L-type intrinsic Ca(2+)- currents, inhibitory synapses help in maintaining the slow depolarization, although they accomplish the role of depressing firing at the beginning of the response. We then used a NEURON model of spiny cells to show that inhibitory synapses arriving asynchronously on the dendrites can help to simulate a plateau potential similar to that observed experimentally.
Figures




Similar articles
-
Spontaneous voltage oscillations in striatal projection neurons in a rat corticostriatal slice.J Physiol. 2003 Nov 15;553(Pt 1):169-82. doi: 10.1113/jphysiol.2003.050799. Epub 2003 Sep 8. J Physiol. 2003. PMID: 12963790 Free PMC article.
-
Glutamatergic and GABAergic postsynaptic responses of striatal spiny neurons to intrastriatal and cortical stimulation recorded in slice preparations.Neuroscience. 1996 Feb;70(4):925-40. doi: 10.1016/0306-4522(95)00410-6. Neuroscience. 1996. PMID: 8848174
-
Contribution of different classes of glutamate receptors in the corticostriatal polysynaptic responses from striatal direct and indirect projection neurons.BMC Neurosci. 2013 Jun 20;14:60. doi: 10.1186/1471-2202-14-60. BMC Neurosci. 2013. PMID: 23782743 Free PMC article.
-
Functional diversity and specificity of neostriatal interneurons.Curr Opin Neurobiol. 2004 Dec;14(6):685-92. doi: 10.1016/j.conb.2004.10.003. Curr Opin Neurobiol. 2004. PMID: 15582369 Review.
-
Role of giant depolarizing potentials in shaping synaptic currents in the developing hippocampus.Crit Rev Neurobiol. 2006;18(1-2):13-23. doi: 10.1615/critrevneurobiol.v18.i1-2.30. Crit Rev Neurobiol. 2006. PMID: 17725505 Review.
Cited by
-
Dopaminergic modulation of corticostriatal responses in medium spiny projection neurons from direct and indirect pathways.Front Syst Neurosci. 2011 Mar 29;5:15. doi: 10.3389/fnsys.2011.00015. eCollection 2011. Front Syst Neurosci. 2011. PMID: 21483724 Free PMC article.
-
Duration differences of corticostriatal responses in striatal projection neurons depend on calcium activated potassium currents.Front Syst Neurosci. 2013 Oct 4;7:63. doi: 10.3389/fnsys.2013.00063. eCollection 2013. Front Syst Neurosci. 2013. PMID: 24109439 Free PMC article.
-
Dynamic modulation of spike timing-dependent calcium influx during corticostriatal upstates.J Neurophysiol. 2013 Oct;110(7):1631-45. doi: 10.1152/jn.00232.2013. Epub 2013 Jul 10. J Neurophysiol. 2013. PMID: 23843436 Free PMC article.
-
Different corticostriatal integration in spiny projection neurons from direct and indirect pathways.Front Syst Neurosci. 2010 Jun 10;4:15. doi: 10.3389/fnsys.2010.00015. eCollection 2010. Front Syst Neurosci. 2010. PMID: 20589098 Free PMC article.
-
The balance of striatal feedback transmission is disrupted in a model of parkinsonism.J Neurosci. 2013 Mar 13;33(11):4964-75. doi: 10.1523/JNEUROSCI.4721-12.2013. J Neurosci. 2013. PMID: 23486967 Free PMC article.
References
-
- Akopian G, Walsh JP (2002) Corticostriatal paired-pulse potentiation produced by voltage-dependent activation of NMDA receptors and L-type Ca2+ channels. J Neurophysiol 87:157–165 - PubMed
-
- Arshavsky YI, Deliagina TG, Orlovsky GN (1997) Pattern generation. Curr Opin Neurobiol 7:781–789. doi:10.1016/S0959-4388(97)80136-5 - PubMed
-
- Bargas J, Galarraga E, Aceves J (1991) Dendritic activity on neostriatal neurons as inferred from somatic intracellular recordings. Brain Res 539:159–163. doi:10.1016/0006-8993(91)90700-6 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
