

Use of (32)P to study dynamics of the mitochondrial phosphoproteome
- PMID: 19351177
- PMCID: PMC3177856
- DOI: 10.1021/pr800913j
Use of (32)P to study dynamics of the mitochondrial phosphoproteome
Abstract
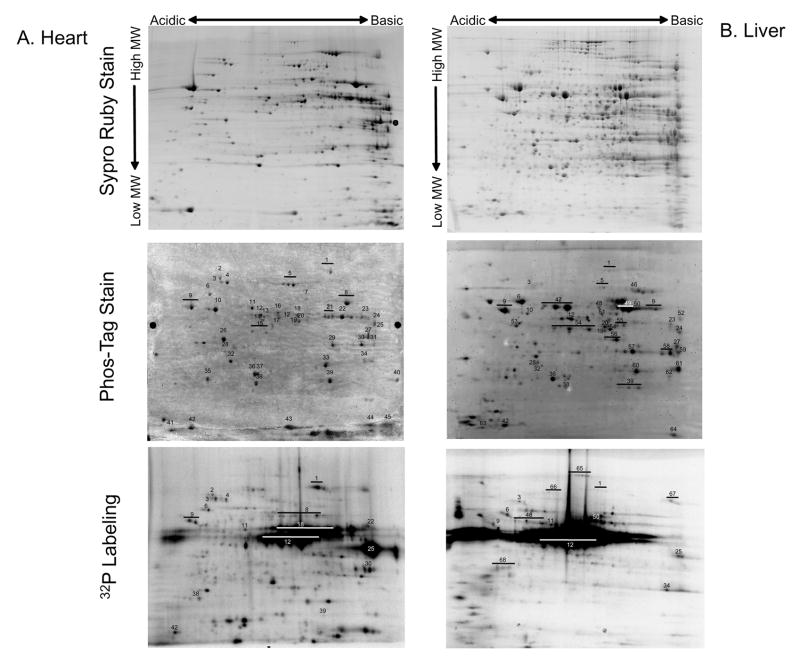
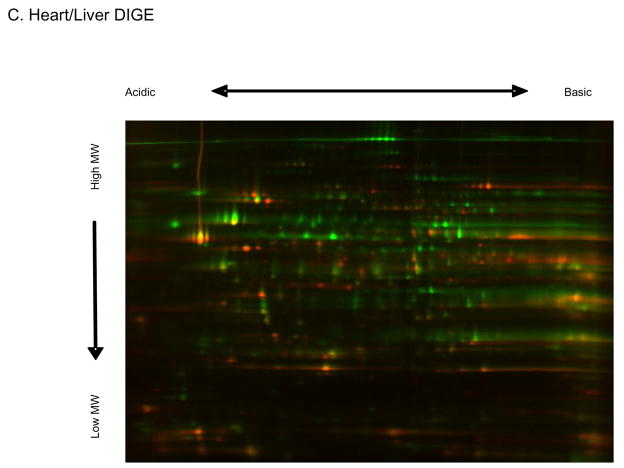
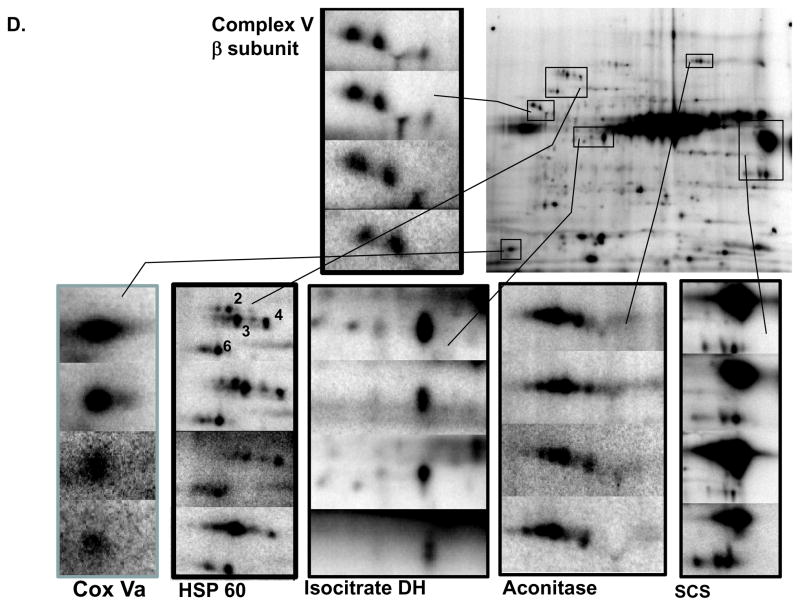
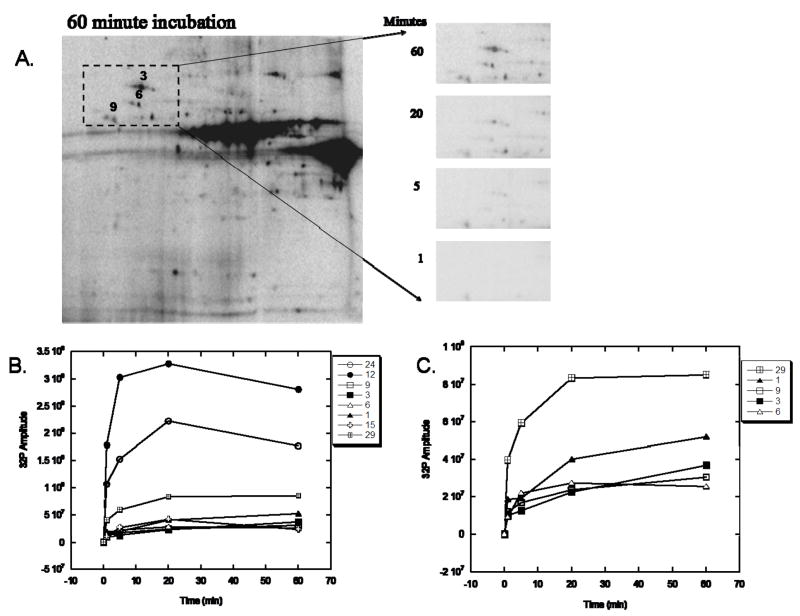
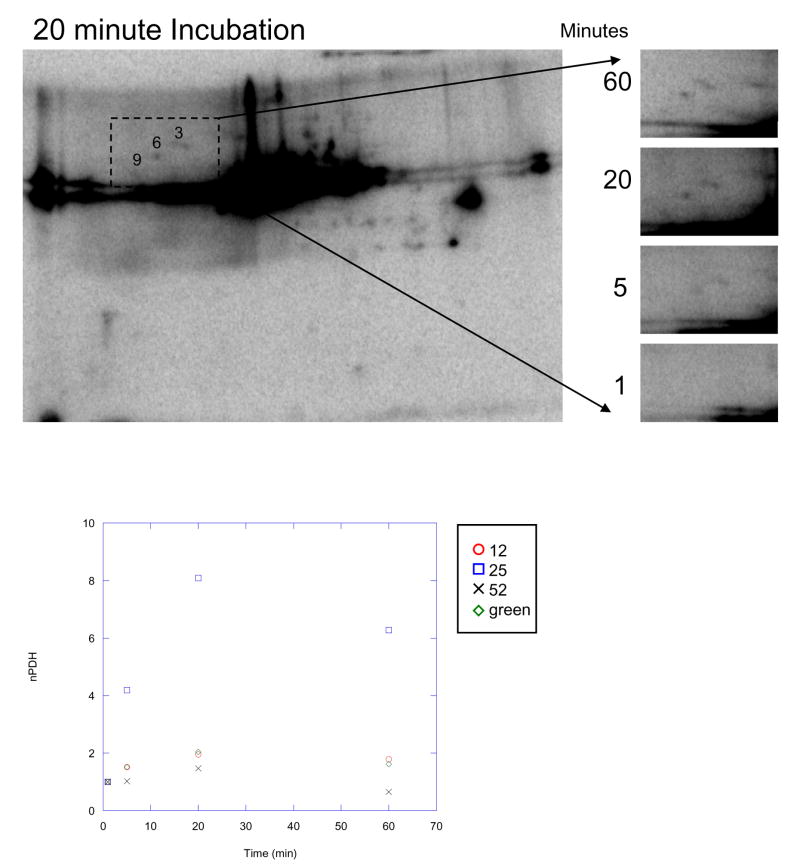
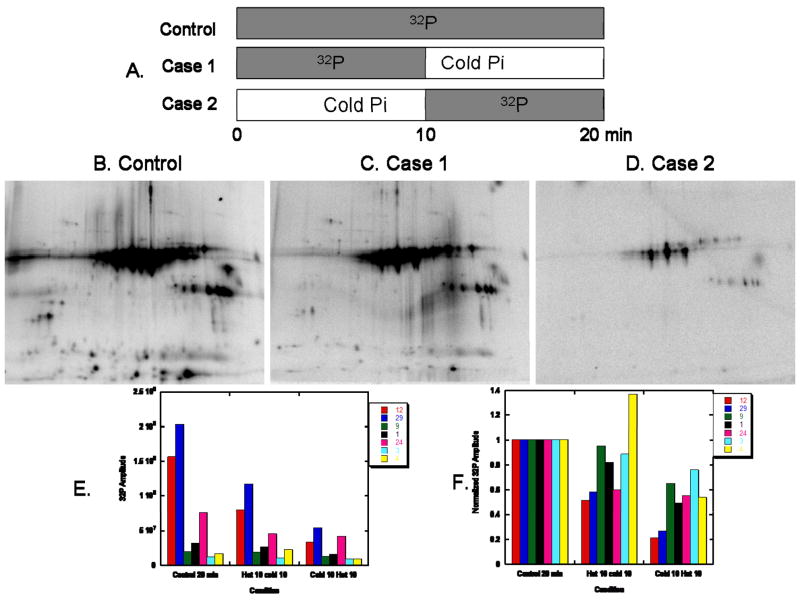
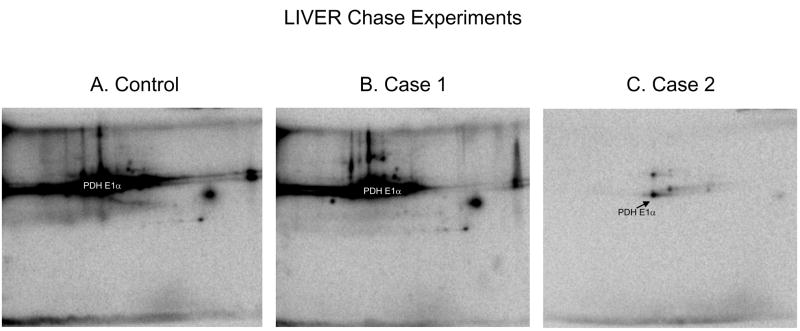
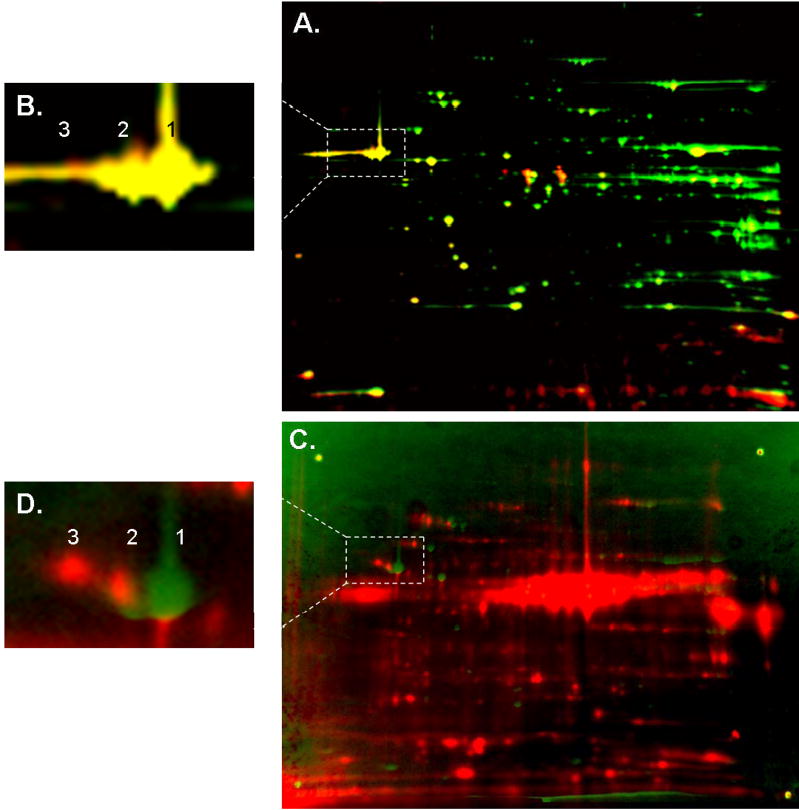
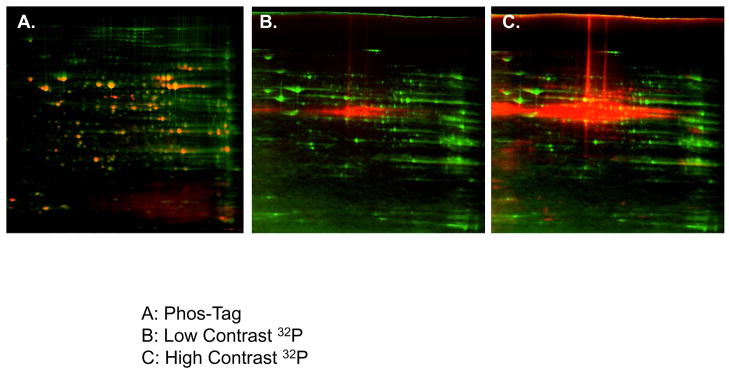
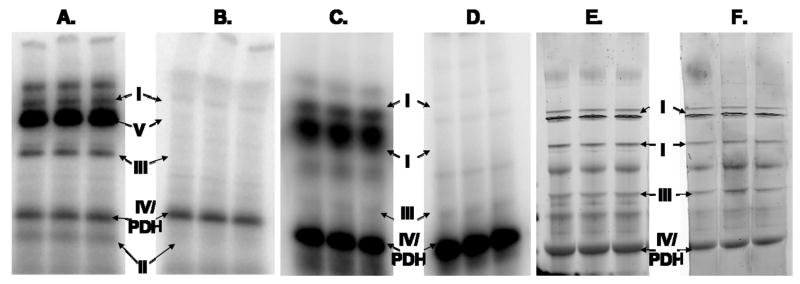
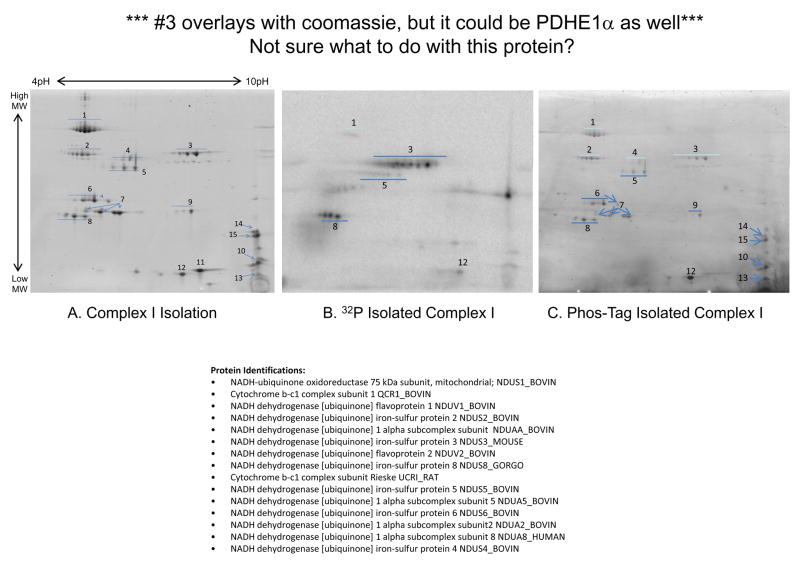
Protein phosphorylation is a well-characterized regulatory mechanism in the cytosol, but remains poorly defined in the mitochondrion. In this study, we characterized the use of (32)P-labeling to monitor the turnover of protein phosphorylation in the heart and liver mitochondria matrix. The (32)P labeling technique was compared and contrasted to Phos-tag protein phosphorylation fluorescent stain and 2D isoelectric focusing. Of the 64 proteins identified by MS spectroscopy in the Phos-Tag gels, over 20 proteins were correlated with (32)P labeling. The high sensitivity of (32)P incorporation detected proteins well below the mass spectrometry and even 2D gel protein detection limits. Phosphate-chase experiments revealed both turnover and phosphate associated protein pool size alterations dependent on initial incubation conditions. Extensive weak phosphate/phosphate metabolite interactions were observed using nondisruptive native gels, providing a novel approach to screen for potential allosteric interactions of phosphate metabolites with matrix proteins. We confirmed the phosphate associations in Complexes V and I due to their critical role in oxidative phosphorylation and to validate the 2D methods. These complexes were isolated by immunocapture, after (32)P labeling in the intact mitochondria, and revealed (32)P-incorporation for the alpha, beta, gamma, OSCP, and d subunits in Complex V and the 75, 51, 42, 23, and 13a kDa subunits in Complex I. These results demonstrate that a dynamic and extensive mitochondrial matrix phosphoproteome exists in heart and liver.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
