

The intracellular II-III loops of Cav1.2 and Cav1.3 uncouple L-type voltage-gated Ca2+ channels from glucagon-like peptide-1 potentiation of insulin secretion in INS-1 cells via displacement from lipid rafts
- PMID: 19351867
- PMCID: PMC2700170
- DOI: 10.1124/jpet.109.150672
The intracellular II-III loops of Cav1.2 and Cav1.3 uncouple L-type voltage-gated Ca2+ channels from glucagon-like peptide-1 potentiation of insulin secretion in INS-1 cells via displacement from lipid rafts
Abstract
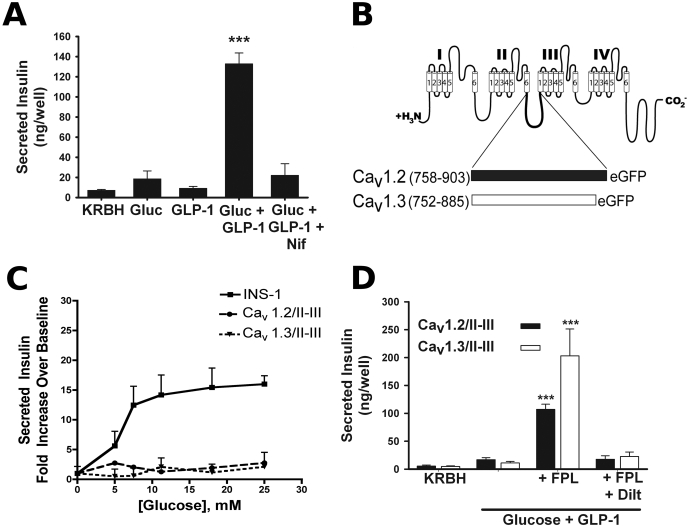
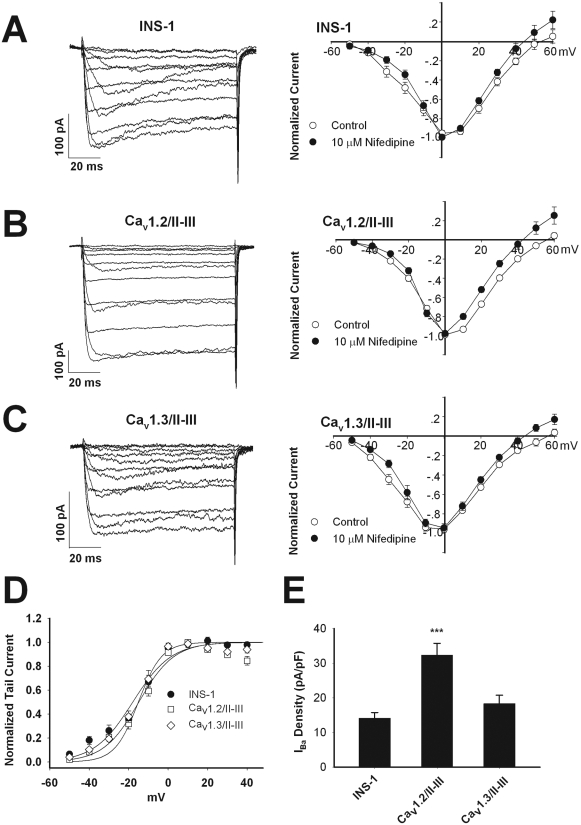
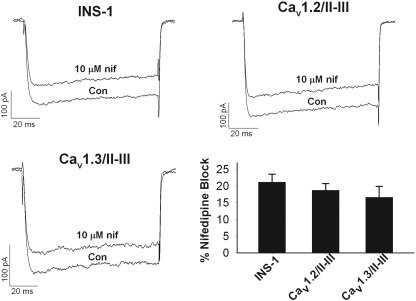
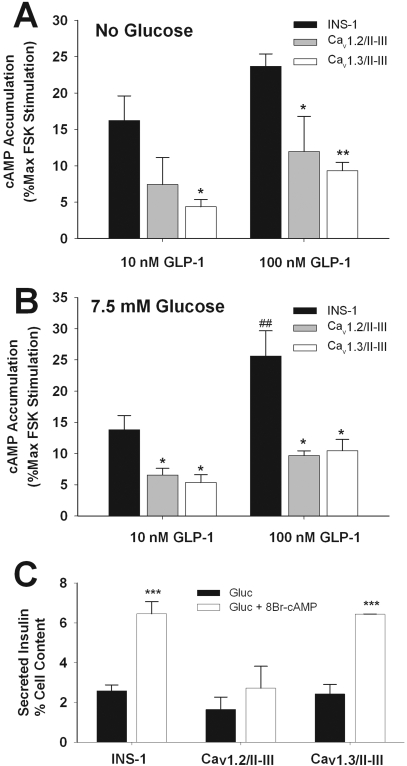
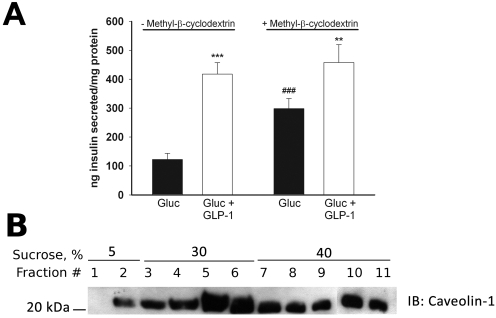
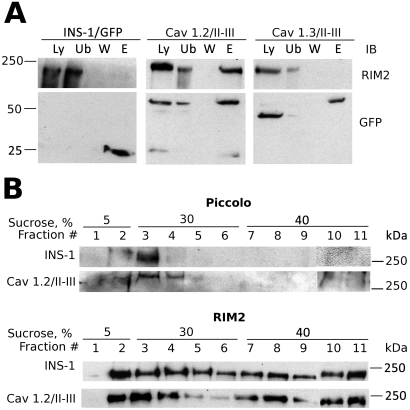
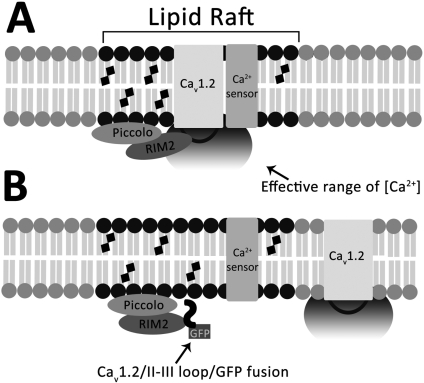
L-type Ca(2+) channels play a key role in the integration of physiological signals regulating insulin secretion that probably requires their localization to specific subdomains of the plasma membrane. We investigated the role of the intracellular II-III loop domains of the L-type channels Ca(v)1.2 and 1.3 in coupling of Ca(2+) influx with glucose-stimulated insulin secretion (GSIS) potentiated by the incretin hormone glucagon-like peptide (GLP)-1. In INS-1 cell lines expressing the Ca(v)1.2/II-III or Ca(v)1.3/II-III peptides, GLP-1 potentiation of GSIS was inhibited markedly, coincident with a decrease in GLP-1-stimulated cAMP accumulation and the redistribution of Ca(v)1.2 and Ca(v)1.3 out of lipid rafts. Neither the Ca(v)1.2/II-III nor the Ca(v)1.3/II-III peptide decreased L-type current density compared with untransfected INS-1 cells. GLP-1 potentiation of GSIS was restored by the L-type channel agonist 2,5-dimethyl-4-[2-(phenylmethyl)benzoyl]-1H-pyrrole-3-carboxylic acid methyl ester (FPL-64176). In contrast, potentiation of GSIS by 8-bromo-cAMP (8-Br-cAMP) was inhibited in Ca(v)1.2/II-III but not Ca(v)1.3/II-III cells. These differences may involve unique protein-protein interactions because the Ca(v)1.2/II-III peptide, but not the Ca(v)1.3/II-III peptide, immunoprecipitates Rab3-interacting molecule (RIM) 2 from INS-1 cell lysates. RIM2, and its binding partner Piccolo, localize to lipid rafts, and they may serve as anchors for Ca(v)1.2 localization to lipid rafts in INS-1 cells. These findings suggest that the II-III interdomain loops of Ca(v)1.2, and possibly Ca(v)1.3, direct these channels to membrane microdomains in which the proteins that mediate potentiation of GSIS by GLP-1 and 8-Br-cAMP assemble.
Figures

References
-
- Asfari M, Janjic D, Meda P, Li G, Halban PA, and Wollheim CB (1992) Establishment of 2-mercaptoethanol-dependent differentiated insulin-secreting cell lines. Endocrinology 130 167-178. - PubMed
-
- Bergsten P (2002) Role of oscillations in membrane potential, cytoplasmic Ca2+, and metabolism for plasma insulin oscillations. Diabetes 51(Suppl 1): S171-S176. - PubMed
-
- Catterall WA (1995) Structure and function of voltage-gated ion channels. Annu Rev Biochem 64 493-531. - PubMed
-
- Cohen AW, Hnasko R, Schubert W, and Lisanti MP (2004) Role of caveolae and caveolins in health and disease. Physiol Rev 84 1341-1379. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
