

Peroxidase mechanism of lipid-dependent cross-linking of synuclein with cytochrome C: protection against apoptosis versus delayed oxidative stress in Parkinson disease
- PMID: 19351880
- PMCID: PMC2708890
- DOI: 10.1074/jbc.M900418200
Peroxidase mechanism of lipid-dependent cross-linking of synuclein with cytochrome C: protection against apoptosis versus delayed oxidative stress in Parkinson disease
Abstract
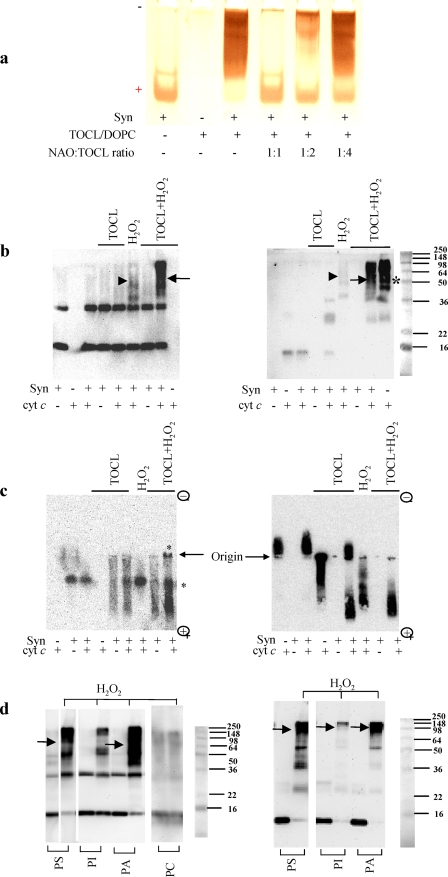
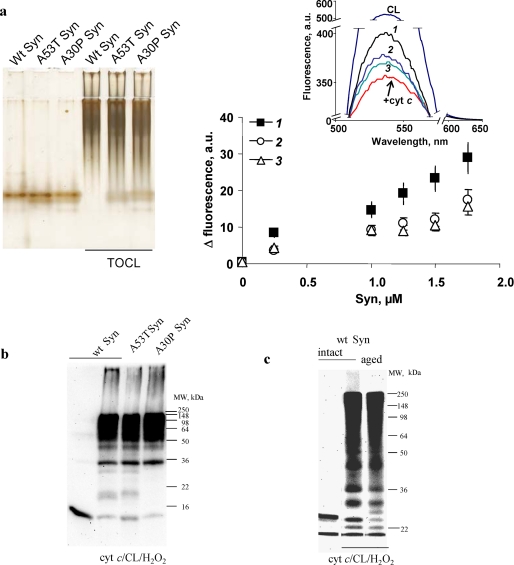
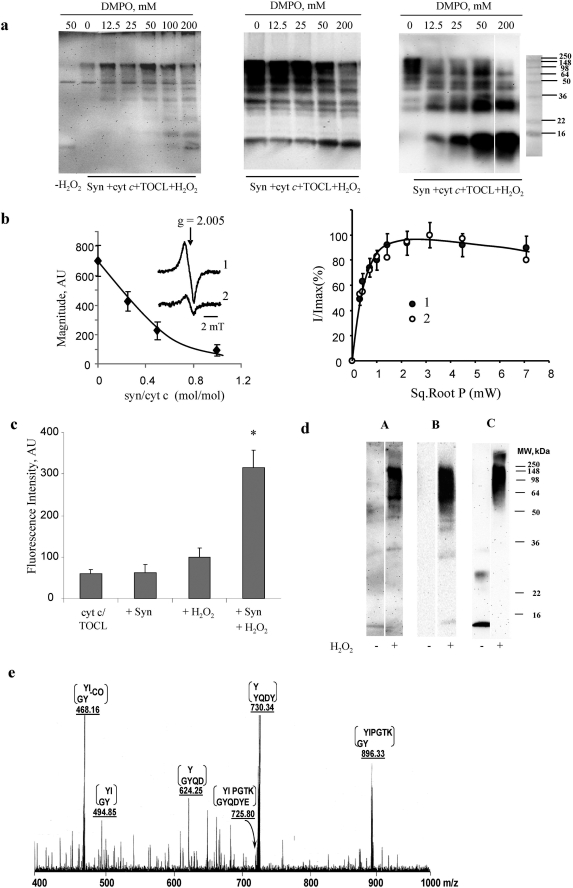
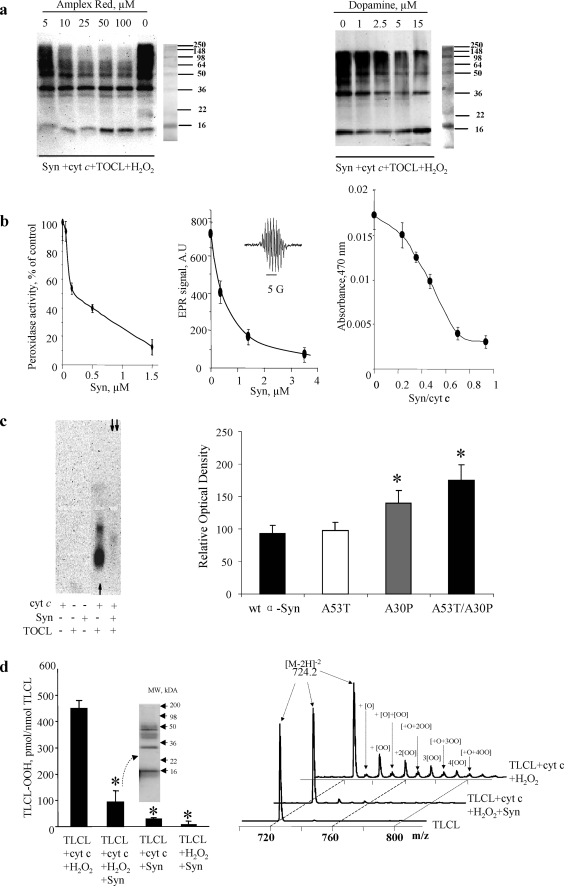
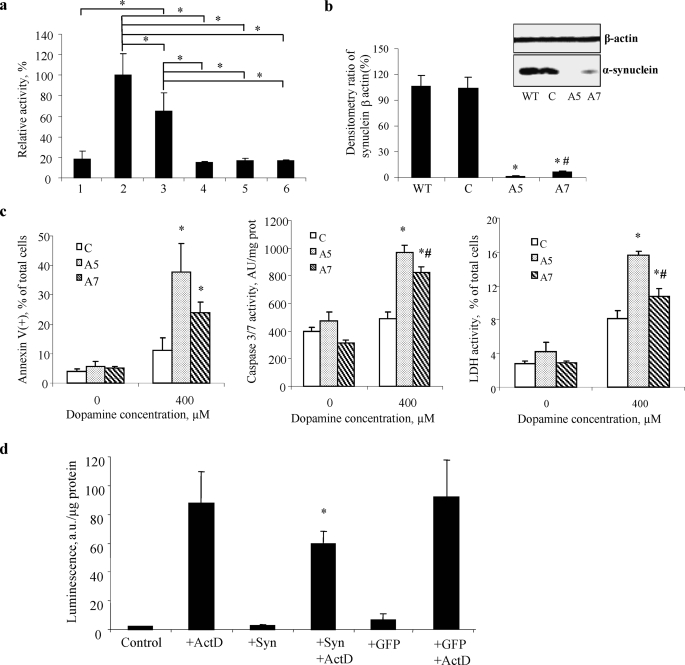
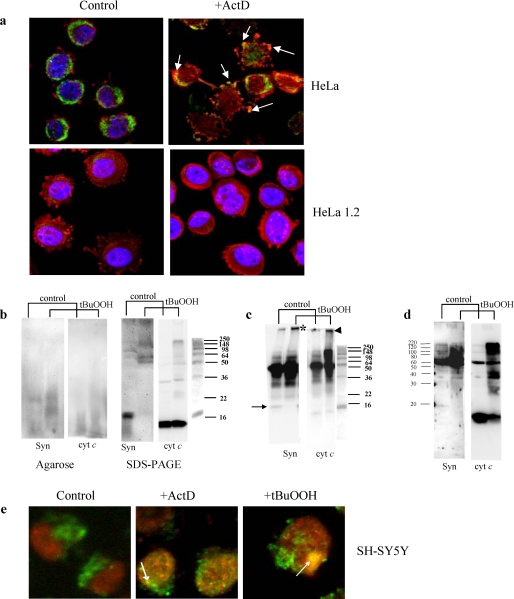
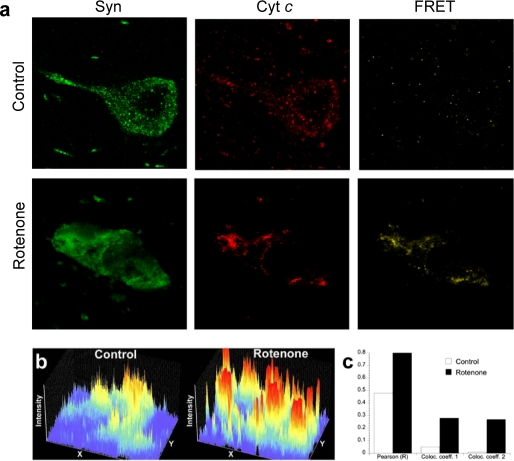
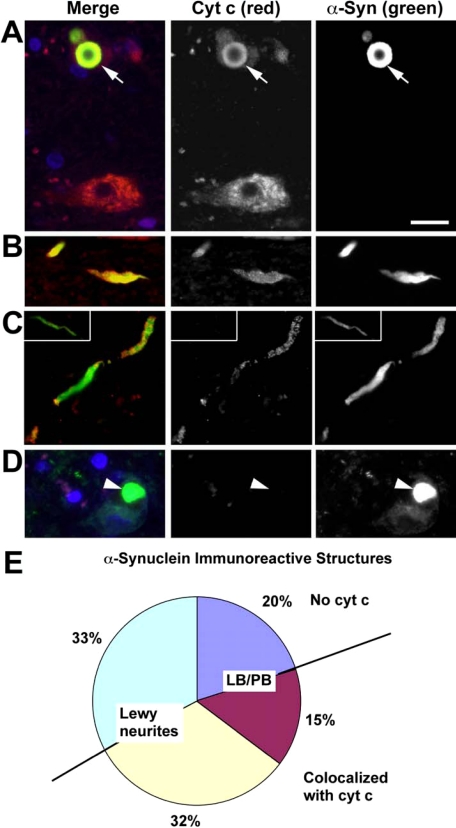
Damage of presynaptic mitochondria could result in release of proapoptotic factors that threaten the integrity of the entire neuron. We discovered that alpha-synuclein (Syn) forms a triple complex with anionic lipids (such as cardiolipin) and cytochrome c, which exerts a peroxidase activity. The latter catalyzes covalent hetero-oligomerization of Syn with cytochrome c into high molecular weight aggregates. Syn is a preferred substrate of this reaction and is oxidized more readily than cardiolipin, dopamine, and other phenolic substrates. Co-localization of Syn with cytochrome c was detected in aggregates formed upon proapoptotic stimulation of SH-SY5Y and HeLa cells and in dopaminergic substantia nigra neurons of rotenone-treated rats. Syn-cardiolipin exerted protection against cytochrome c-induced caspase-3 activation in a cell-free system, particularly in the presence of H(2)O(2). Direct delivery of Syn into mouse embryonic cells conferred resistance to proapoptotic caspase-3 activation. Conversely, small interfering RNA depletion of Syn in HeLa cells made them more sensitive to dopamine-induced apoptosis. In human Parkinson disease substantia nigra neurons, two-thirds of co-localized Syn-cytochrome c complexes occurred in Lewy neurites. Taken together, these results indicate that Syn may prevent execution of apoptosis in neurons through covalent hetero-oligomerization of cytochrome c. This immediate protective function of Syn is associated with the formation of the peroxidase complex representing a source of oxidative stress and postponed damage.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- NS061817/NS/NINDS NIH HHS/United States
- AG026389/AG/NIA NIH HHS/United States
- R37 NS061817/NS/NINDS NIH HHS/United States
- R21 HD057587/HD/NICHD NIH HHS/United States
- ES012068/ES/NIEHS NIH HHS/United States
- AG05133/AG/NIA NIH HHS/United States
- R01 NS061817/NS/NINDS NIH HHS/United States
- P50 AG005133/AG/NIA NIH HHS/United States
- R01 AG026389/AG/NIA NIH HHS/United States
- U54 ES012068/ES/NIEHS NIH HHS/United States
- P01 NS059806/NS/NINDS NIH HHS/United States
- U19 AIO68021/PHS HHS/United States
- U19 AI068021/AI/NIAID NIH HHS/United States
- HD057587/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
