

Spatiotemporal patterning during T cell activation is highly diverse
- PMID: 19351954
- PMCID: PMC2694444
- DOI: 10.1126/scisignal.2000199
Spatiotemporal patterning during T cell activation is highly diverse
Abstract
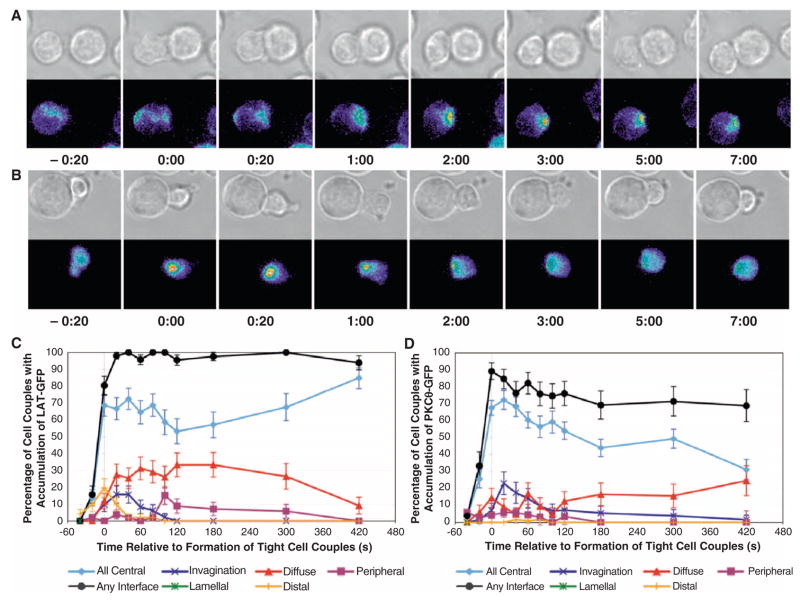
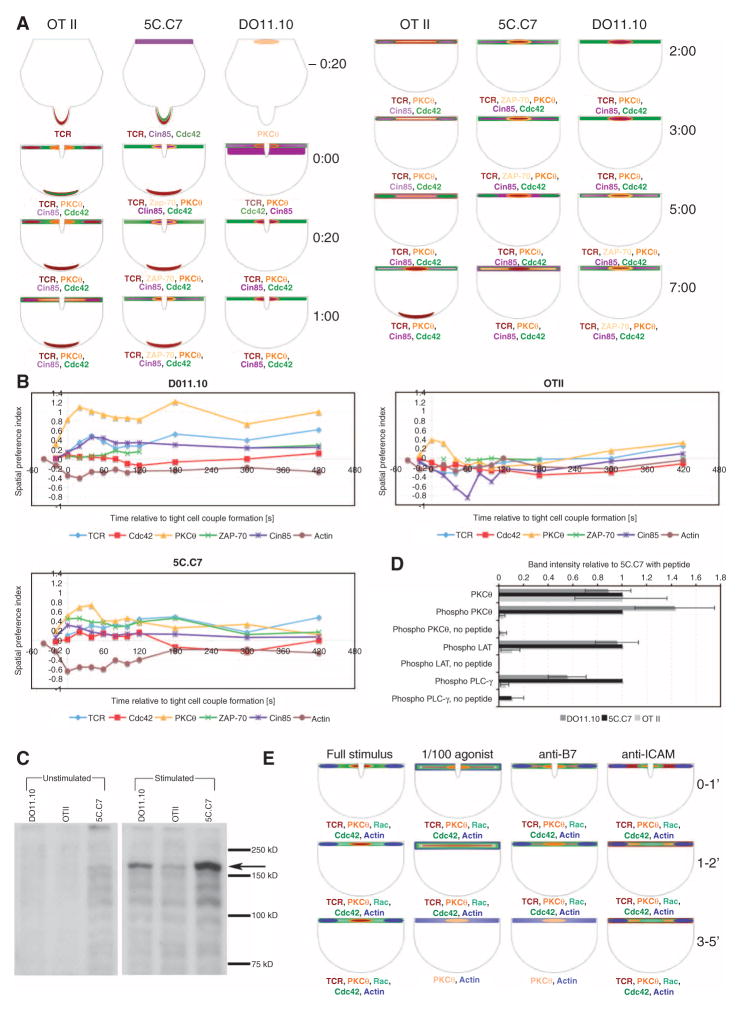
Temporal and spatial variations in the concentrations of signaling intermediates in a living cell are important for signaling in complex networks because they modulate the probabilities that signaling intermediates will interact with each other. We have studied 30 signaling sensors, ranging from receptors to transcription factors, in the physiological activation of murine ex vivo T cells by antigen-presenting cells. Spatiotemporal patterning of these molecules was highly diverse and varied with specific T cell receptors and T cell activation conditions. The diversity and variability observed suggest that spatiotemporal patterning controls signaling interactions during T cell activation in a physiologically important and discriminating manner. In support of this, the effective clustering of a group of ligand-engaged receptors and signaling intermediates in a joint pattern consistently correlated with efficient T cell activation at the level of the whole cell.
Figures



References
-
- Kavran JM, Klein DE, Lee A, Falasca M, Isakoff SJ, Skolnik EY, Lemmon MA. Specificity and promiscuity in phosphoinositide binding by pleckstrin homology domains. J Biol Chem. 1998;273:30497–30508. - PubMed
-
- Songyang Z, Shoelson SE, Chaudhuri M, Gish G, Pawson T, Haser WG, King F, Roberts T, Ratnofsky S, Lechleider RJ, Neel BG, Birge RB, Fajardo JE, Chou MM, Hanafusa H, Schaffhausen B, Cantley LC. SH2 domains recognize specific phosphopeptide sequences. Cell. 1993;72:767–778. - PubMed
-
- Pawson T, Nash P. Assembly of cell regulatory systems through protein interaction domains. Science. 2003;300:445–452. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
