

Parallel processing strategies of the primate visual system
- PMID: 19352403
- PMCID: PMC2771435
- DOI: 10.1038/nrn2619
Parallel processing strategies of the primate visual system
Abstract
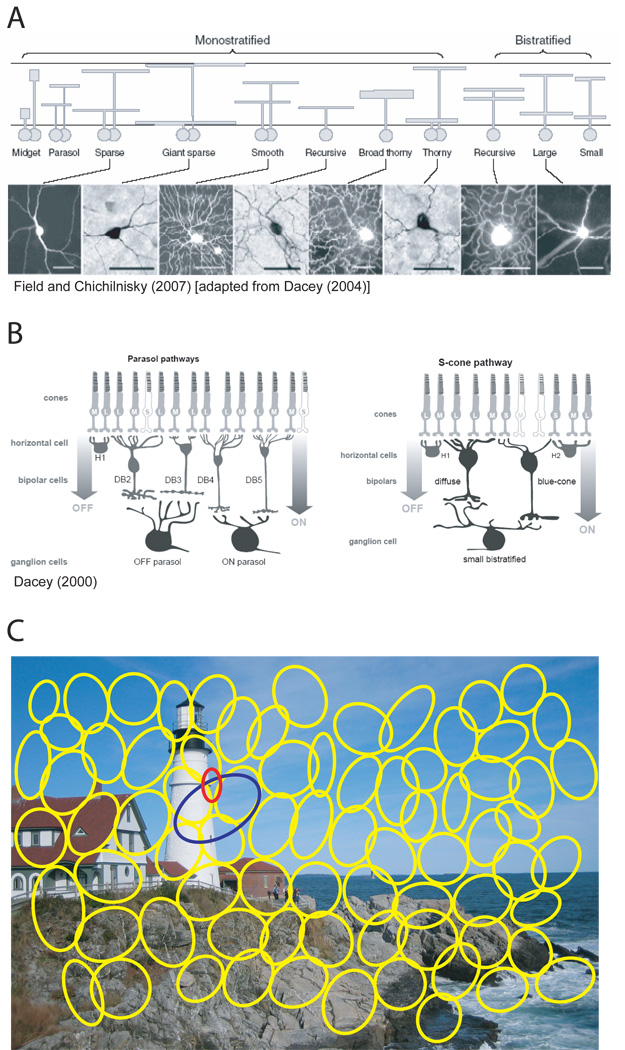
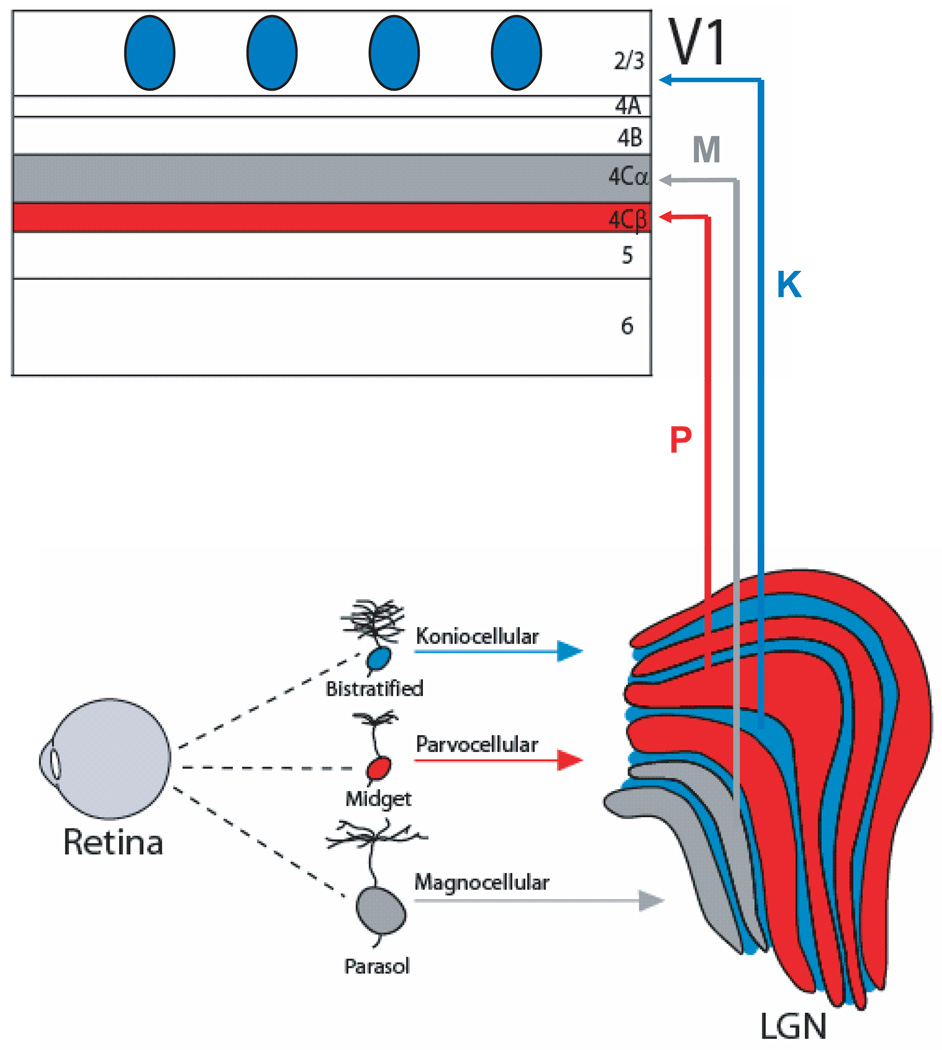
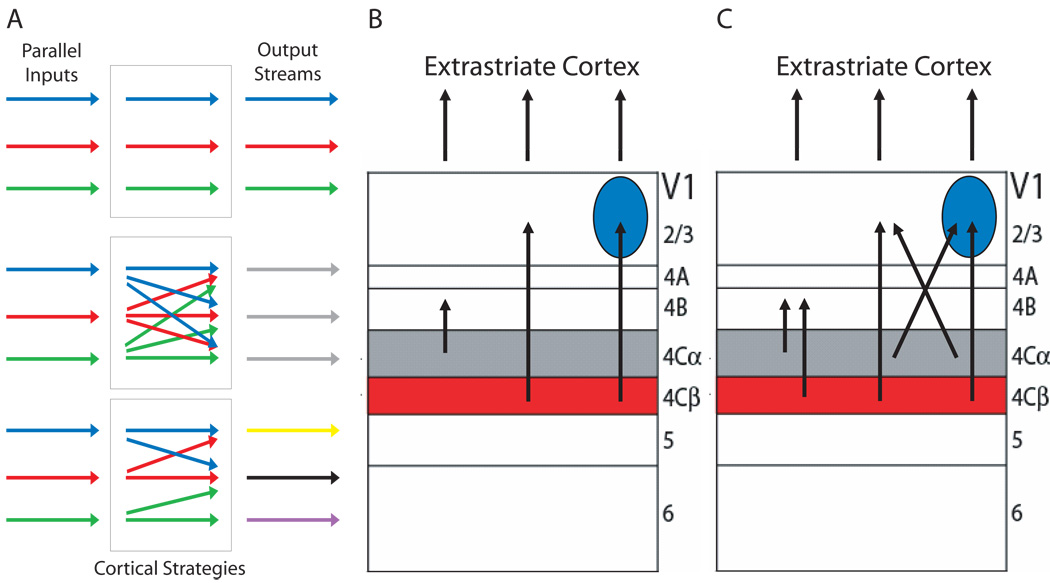
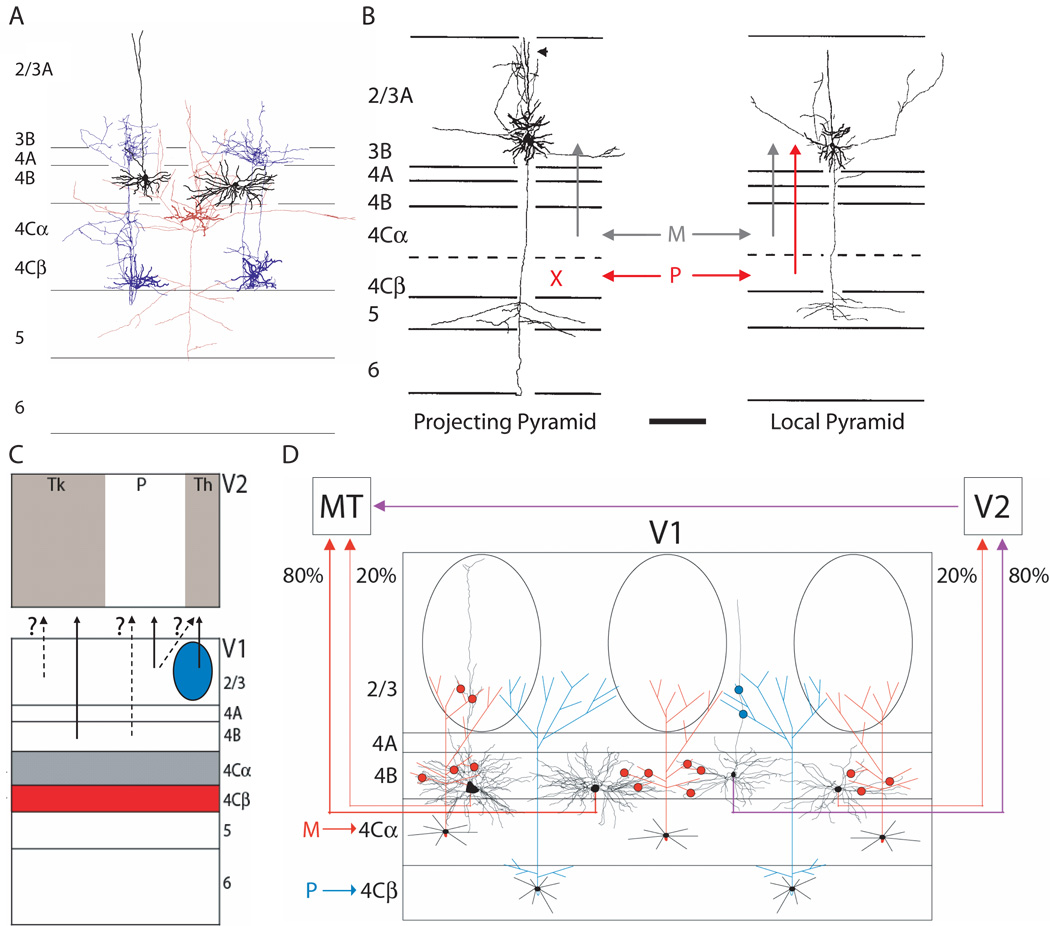
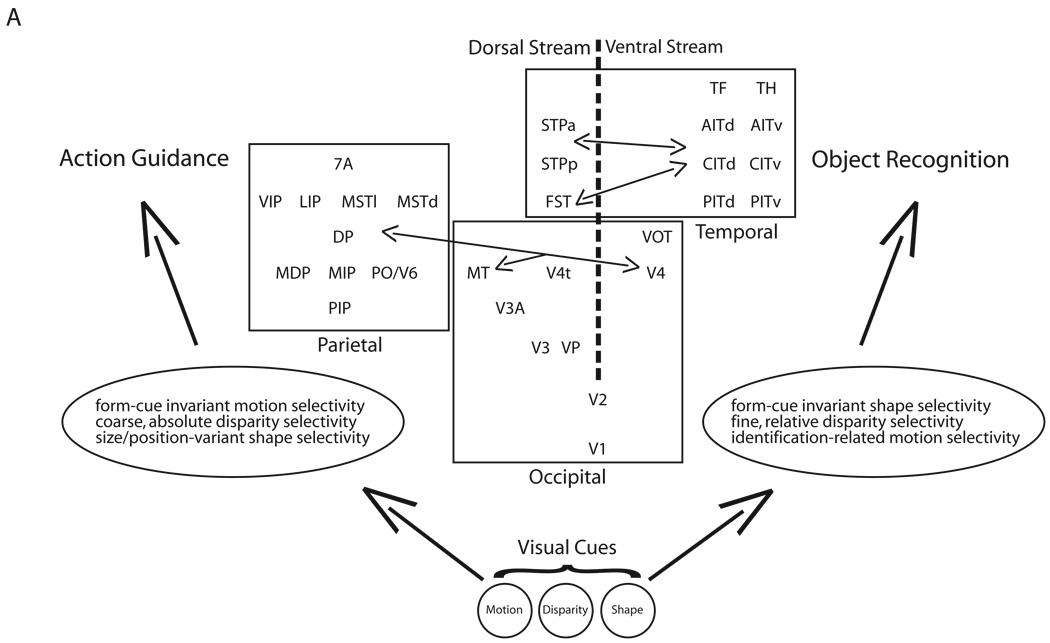
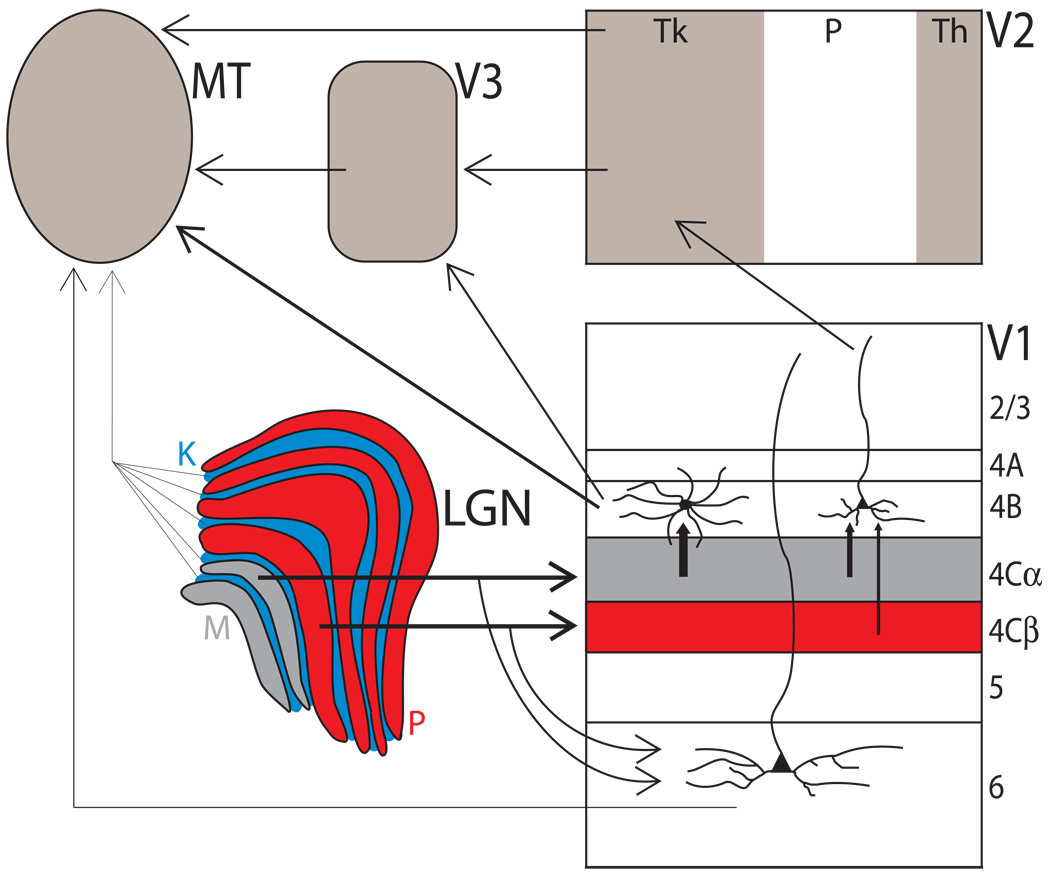
Incoming sensory information is sent to the brain along modality-specific channels corresponding to the five senses. Each of these channels further parses the incoming signals into parallel streams to provide a compact, efficient input to the brain. Ultimately, these parallel input signals must be elaborated on and integrated in the cortex to provide a unified and coherent percept. Recent studies in the primate visual cortex have greatly contributed to our understanding of how this goal is accomplished. Multiple strategies including retinal tiling, hierarchical and parallel processing and modularity, defined spatially and by cell type-specific connectivity, are used by the visual system to recover the intricate detail of our visual surroundings.
Figures
References
-
- Gasser HS, Erlanger J. The role of fiber size in the establishment of a nerve block by pressure or cocaine. Am. J. Physiol. 1929;88:581–591.
-
- Bishop GH. Fiber groups in the optic nerves. Am. J. Physiol. 1933;106:460–470.
-
- Goodale MA, Milner AD. Separate visual pathways for perception and action. Trends Neurosci. 1992;15:20–25. - PubMed
-
- Hendry SH, Reid RC. The koniocellular pathway in primate vision. Annu Rev Neurosci. 2000;23:127–153. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
