

The nucleus accumbens: a switchboard for goal-directed behaviors
- PMID: 19352511
- PMCID: PMC2663037
- DOI: 10.1371/journal.pone.0005062
The nucleus accumbens: a switchboard for goal-directed behaviors
Abstract
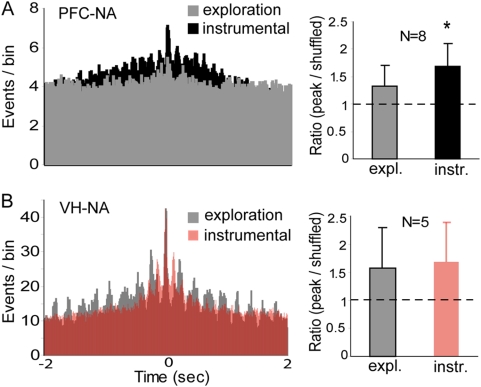
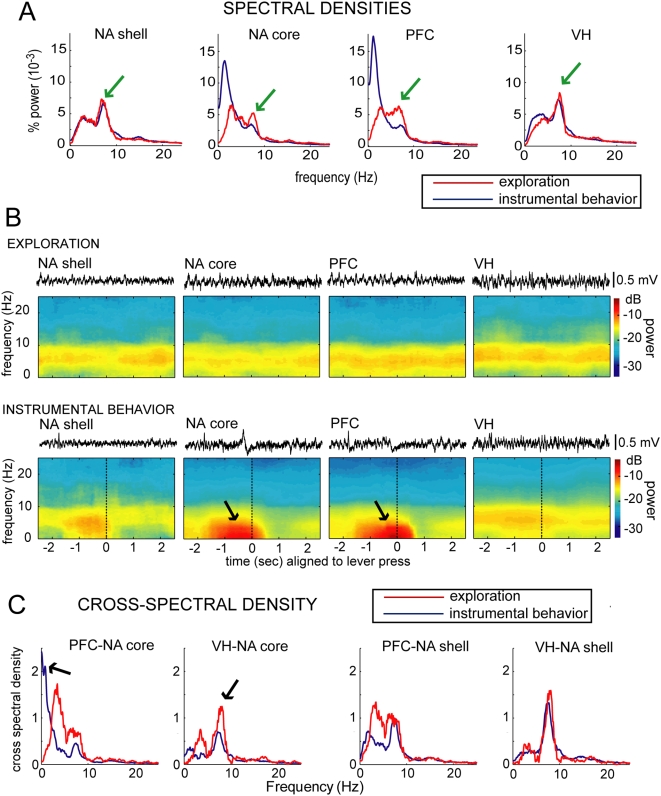
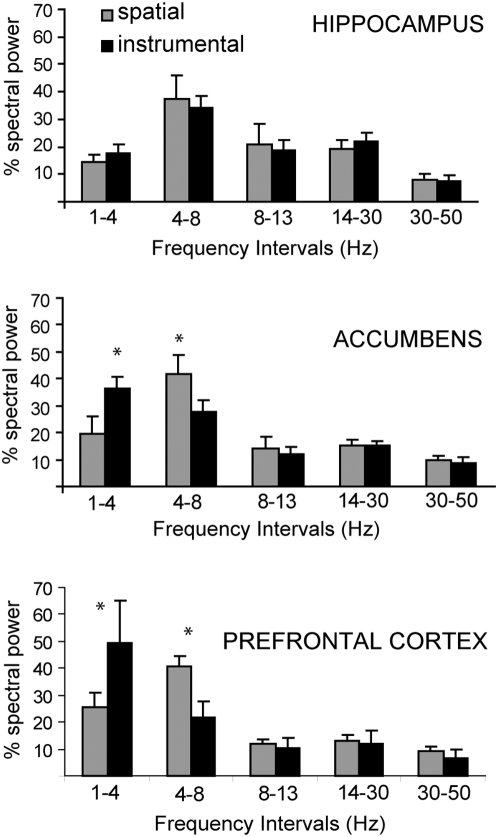
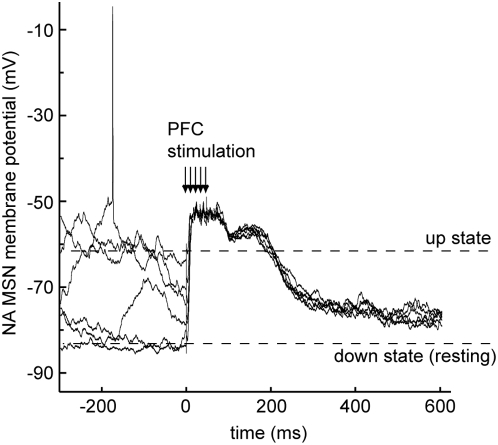
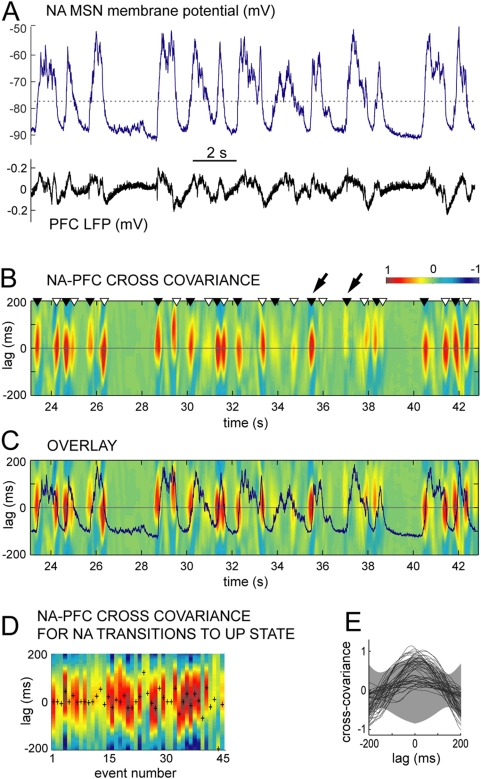
Reward intake optimization requires a balance between exploiting known sources of rewards and exploring for new sources. The prefrontal cortex (PFC) and associated basal ganglia circuits are likely candidates as neural structures responsible for such balance, while the hippocampus may be responsible for spatial/contextual information. Although studies have assessed interactions between hippocampus and PFC, and between hippocampus and the nucleus accumbens (NA), it is not known whether 3-way interactions among these structures vary under different behavioral conditions. Here, we investigated these interactions with multichannel recordings while rats explored an operant chamber and while they performed a learned lever-pressing task for reward in the same chamber shortly afterward. Neural firing and local field potentials in the NA core synchronized with hippocampal activity during spatial exploration, but during lever pressing they instead synchronized more strongly with the PFC. The latter is likely due to transient drive of NA neurons by bursting prefrontal activation, as in vivo intracellular recordings in anesthetized rats revealed that NA up states can transiently synchronize with spontaneous PFC activity and PFC stimulation with a bursting pattern reliably evoked up states in NA neurons. Thus, the ability to switch synchronization in a task-dependent manner indicates that the NA core can dynamically select its inputs to suit environmental demands, thereby contributing to decision-making, a function that was thought to primarily depend on the PFC.
Conflict of interest statement
Figures
References
-
- Fuster JM. The Prefrontal Cortex. Anatomy, Physiology, and Neuropsychology of the frontal lobe. New York: Lippincott-Raven; 1997. p. 333.
-
- Grace AA, Floresco SB, Goto Y, Lodge DJ. Regulation of firing of dopaminergic neurons and control of goal-directed behaviors. Trends Neurosci. 2007;30:220–227. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
