

Mitochondrial peroxiredoxin 3 is more resilient to hyperoxidation than cytoplasmic peroxiredoxins
- PMID: 19356151
- PMCID: PMC3745641
- DOI: 10.1042/BJ20090242
Mitochondrial peroxiredoxin 3 is more resilient to hyperoxidation than cytoplasmic peroxiredoxins
Abstract
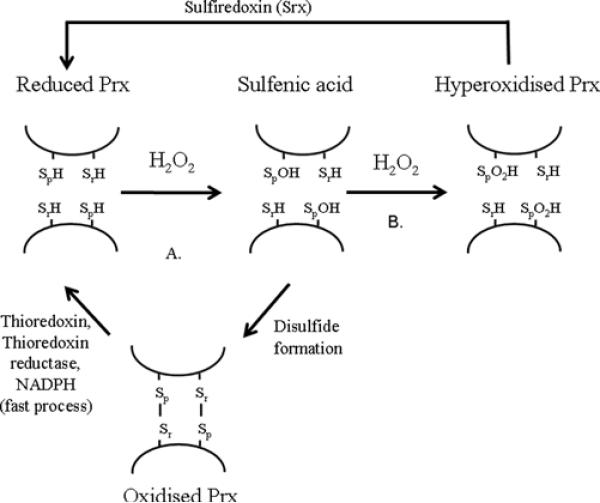
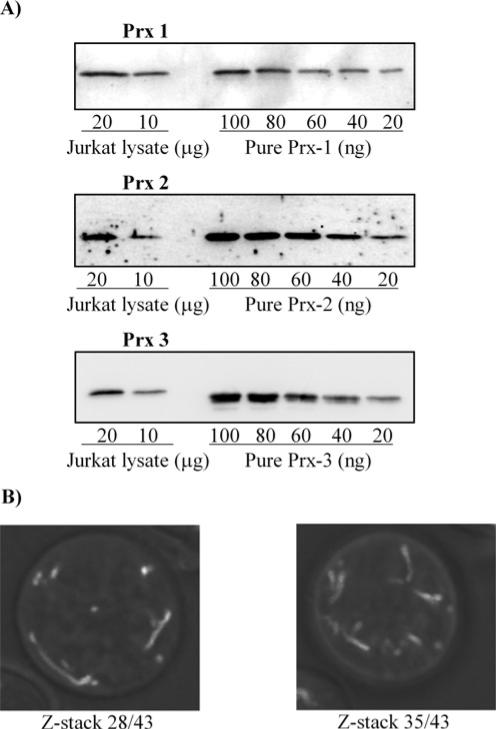
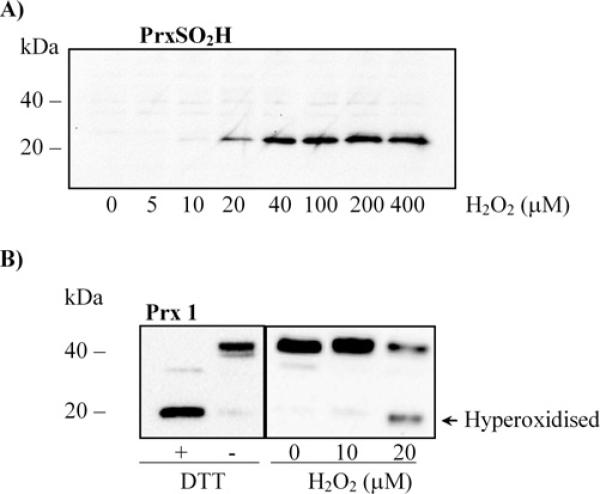
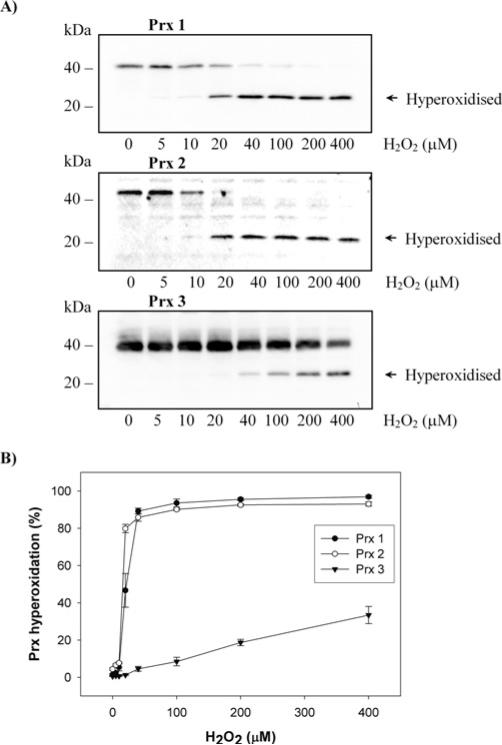
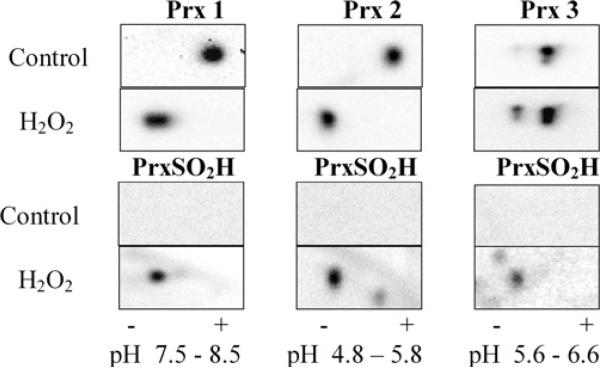
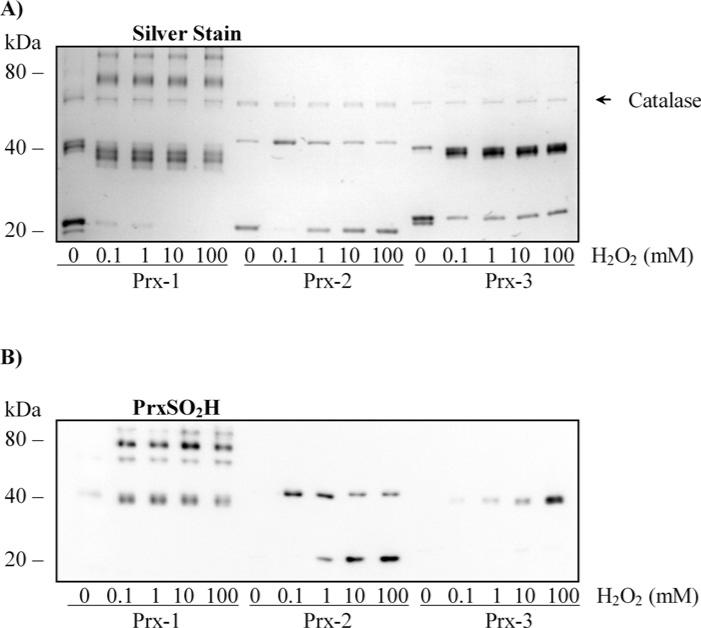
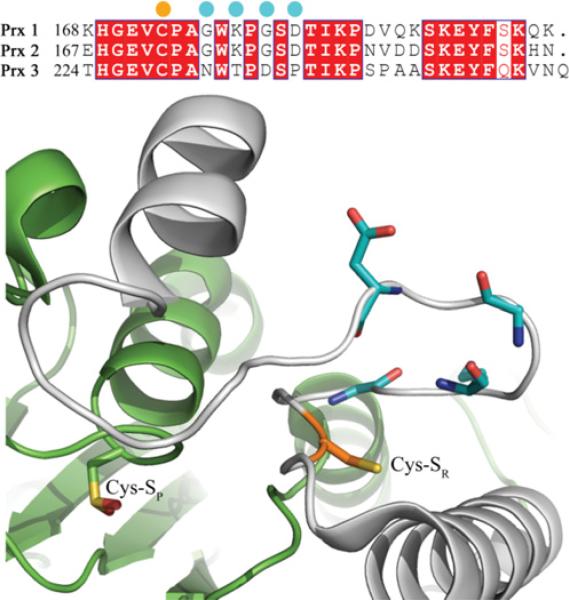
The Prxs (peroxiredoxins) are a family of cysteine-dependent peroxidases that decompose hydrogen peroxide. Prxs become hyperoxidized when a sulfenic acid formed during the catalytic cycle reacts with hydrogen peroxide. In the present study, Western blot methodology was developed to quantify hyperoxidation of individual 2-Cys Prxs in cells. It revealed that Prx 1 and 2 were hyperoxidized at lower doses of hydrogen peroxide than would be predicted from in vitro data, suggesting intracellular factors that promote hyperoxidation. In contrast, mitochondrial Prx 3 was considerably more resistant to hyperoxidation. The concentration of Prx 3 was estimated at 125 microM in the mitochondrial matrix of Jurkat T-lymphoma cells. Although the local cellular environment could influence susceptibility, purified Prx 3 was also more resistant to hyperoxidation, suggesting that despite having C-terminal motifs similar to sensitive eukaryote Prxs, other structural features must contribute to the innate resilience of Prx 3 to hyperoxidation.
Figures

References
-
- Wood ZA, Schroder E, Harris JR, Poole LB. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003;28:32–40. - PubMed
-
- Chae HZ, Chung SJ, Rhee SG. Thioredoxin-dependent peroxide reductase from yeast. J. Biol. Chem. 1994;269:27670–27678. - PubMed
-
- Rabilloud T, Heller M, Gasnier F, Luche S, Rey C, Aebersold R, Benahmed M, Louisot P, Lunardi J. Proteomics analysis of cellular response to oxidative stress – evidence for in vivo overoxidation of peroxiredoxins at their active site. J. Biol. Chem. 2002;277:19396–19401. - PubMed
-
- Chevallet M, Wagner E, Luche S, van Dorsselaer A, Leize-Wagner E, Rabilloud T. Regeneration of peroxiredoxins during recovery after oxidative stress – only some overoxidized peroxiredoxins can be reduced during recovery after oxidative stress. J. Biol. Chem. 2003;278:37146–37153. - PubMed
-
- Yang KS, Kang SW, Woo HA, Hwang SC, Chae HZ, Kim K, Rhee SG. Inactivation of human peroxiredoxin I during catalysis as the result of the oxidation of the catalytic site cysteine to cysteine-sulfinic acid. J. Biol. Chem. 2002;277:38029–38036. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources