

Characterization of the in vitro HIV-1 capsid assembly pathway
- PMID: 19356593
- PMCID: PMC2667805
- DOI: 10.1016/j.jmb.2009.01.058
Characterization of the in vitro HIV-1 capsid assembly pathway
Abstract
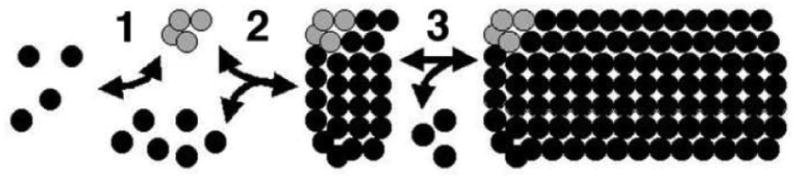
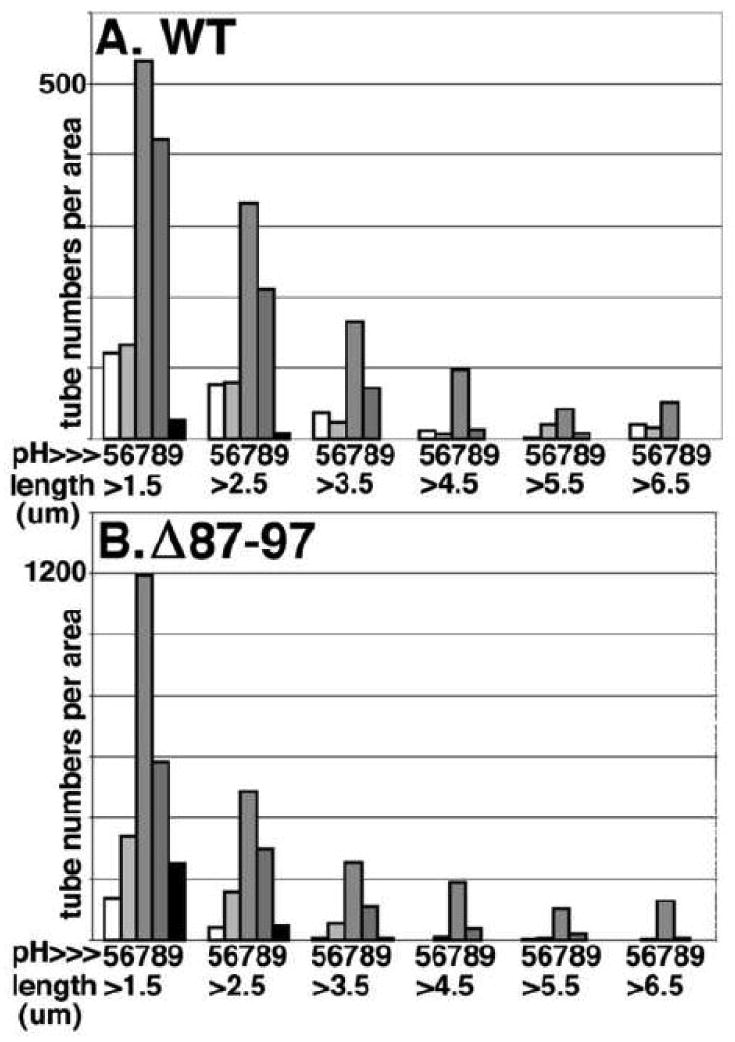
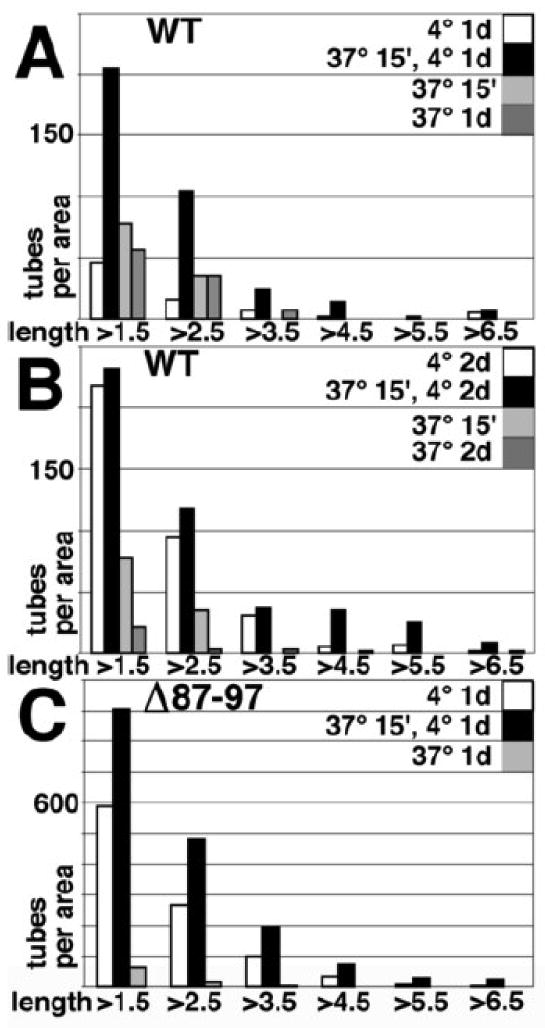
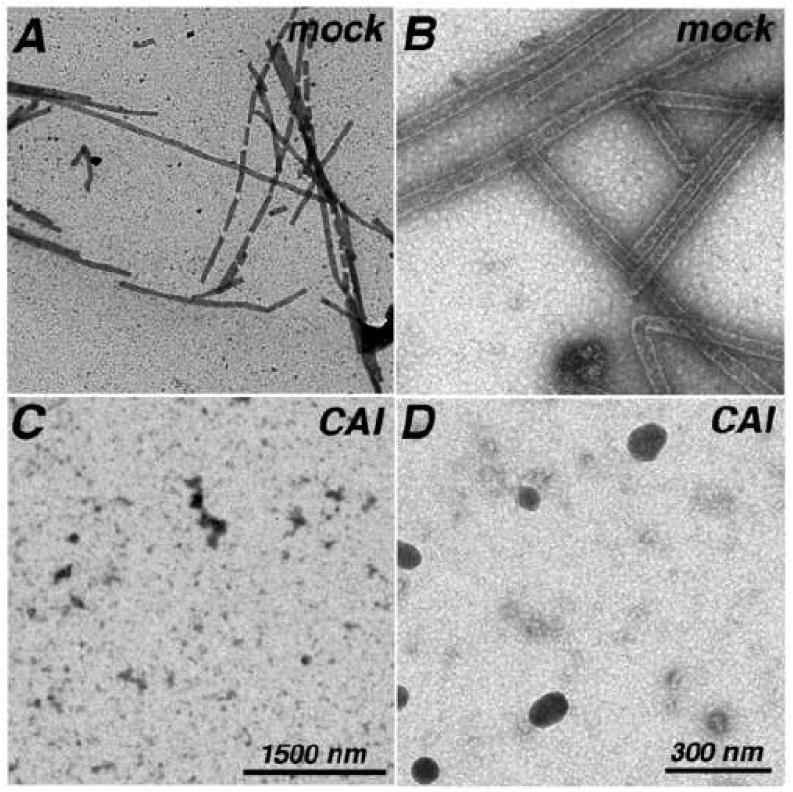
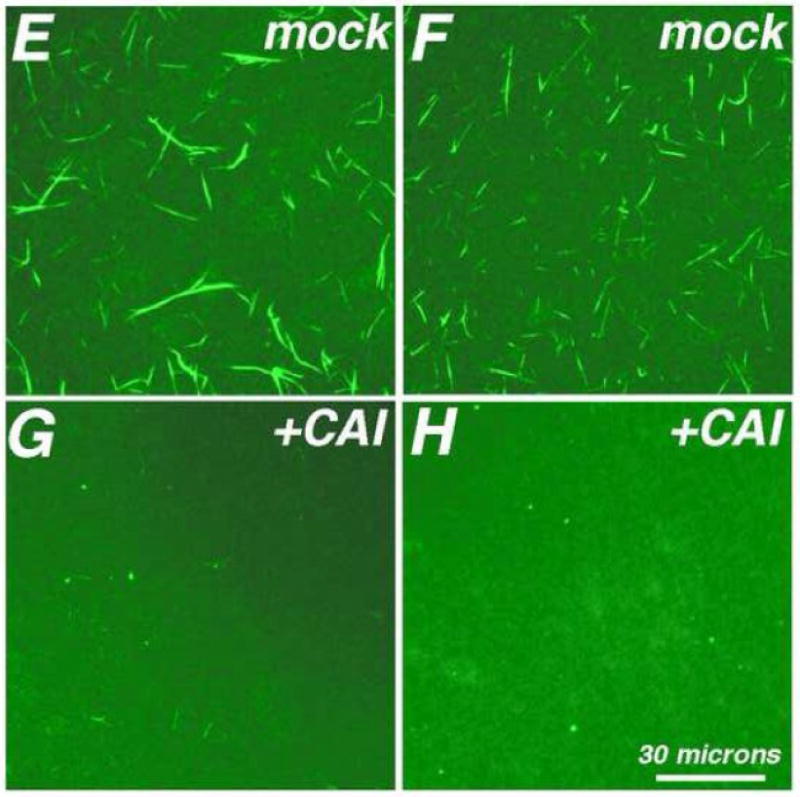
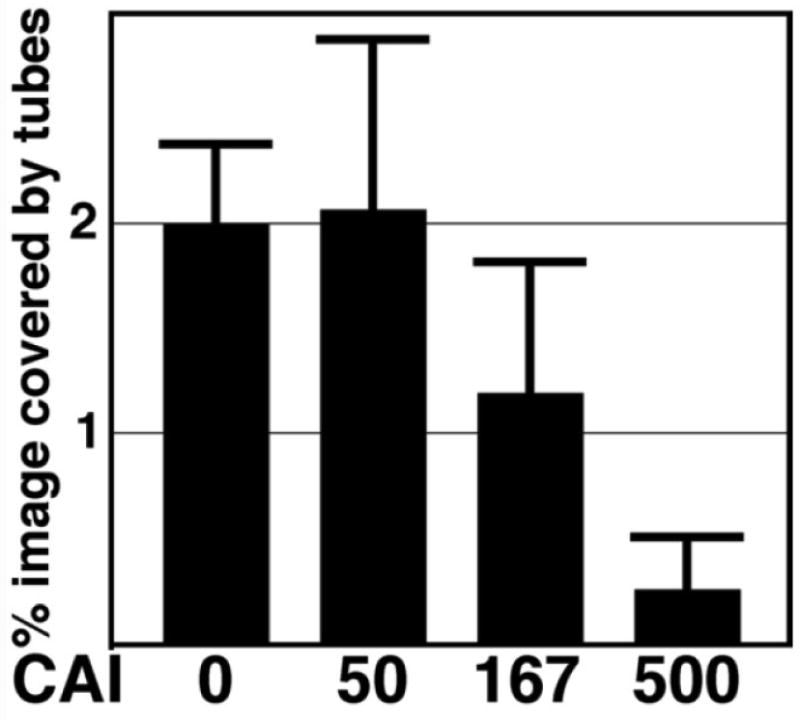
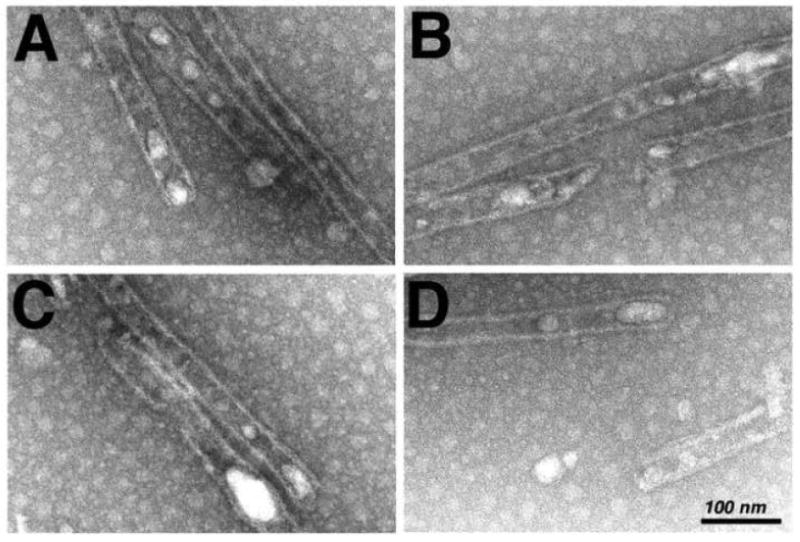
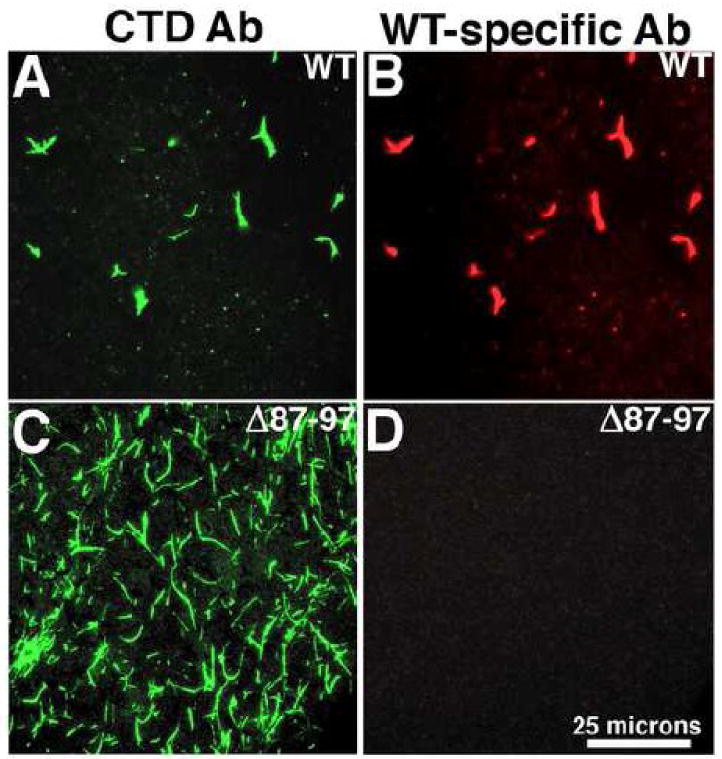
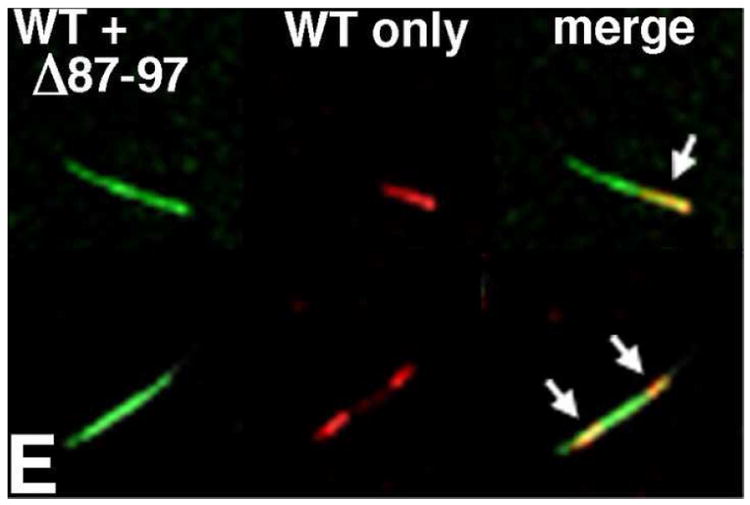
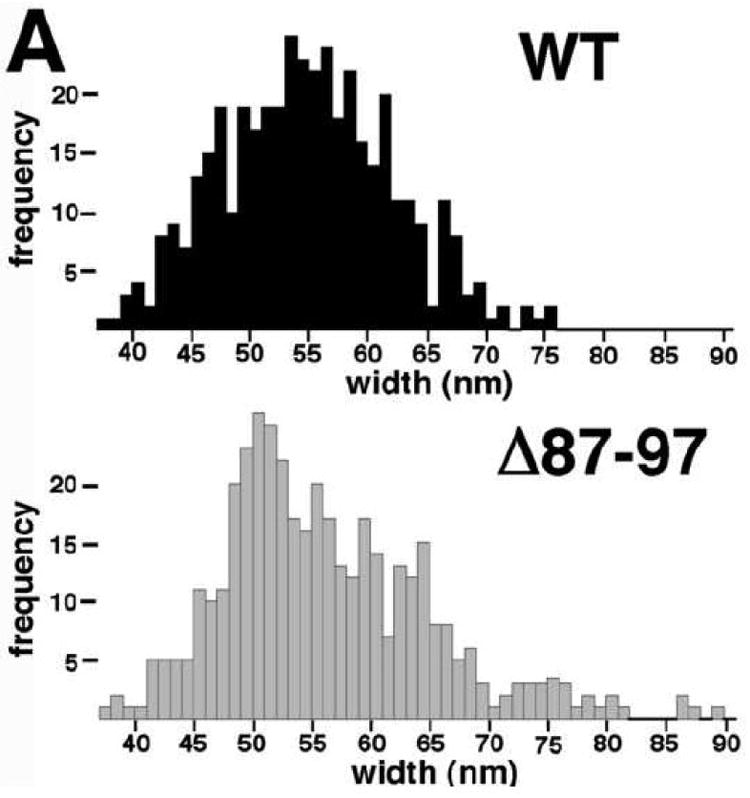
During the morphogenesis of mature human immunodeficiency virus-1 cores, viral capsid proteins assemble conical or tubular shells around viral ribonucleoprotein complexes. This assembly step is mimicked in vitro through reactions in which capsid proteins oligomerize to form long tubes, and this process can be modeled as consisting of a slow nucleation period, followed by a rapid phase of tube growth. We have developed a novel fluorescence microscopy approach to monitor in vitro assembly reactions and have employed it, along with electron microscopy analysis, to characterize the assembly process. Our results indicate that temperature, salt concentration, and pH changes have differential effects on tube nucleation and growth steps. We also demonstrate that assembly can be unidirectional or bidirectional, that growth can be capped, and that proteins can assemble onto the surfaces of tubes, yielding multiwalled or nested structures. Finally, experiments show that a peptide inhibitor of in vitro assembly also can dismantle preexisting tubes, suggesting that such reagents may possess antiviral effects against both viral assembly and uncoating. Our investigations help establish a basis for understanding the mechanism of mature human immunodeficiency virus-1 core assembly and avenues for antiviral inhibition.
Figures







References
-
- Gross I, Hohenberg H, Krausslich HG. In vitro assembly properties of purified bacterially expressed capsid proteins of human immunodeficiency virus. Eur J Biochem. 1997;249:592–600. - PubMed
-
- Swanstrom R, Wills J. Synthesis, assembly and processing of viral proteins. In: Coffin J, Hughes S, Varmus H, editors. Retroviruses. Cold Spring Harbor Laboratory Press; NY: 1997. pp. 263–334. - PubMed
-
- Barklis E, McDermott J, Wilkens S, Fuller S, Thompson D. Organization of HIV-1 capsid proteins on a lipid monolayer. J Biol Chem. 1998;273:7177–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
