

Conditional inactivation of androgen receptor gene in the nervous system: effects on male behavioral and neuroendocrine responses
- PMID: 19357272
- PMCID: PMC6665718
- DOI: 10.1523/JNEUROSCI.0296-09.2009
Conditional inactivation of androgen receptor gene in the nervous system: effects on male behavioral and neuroendocrine responses
Abstract
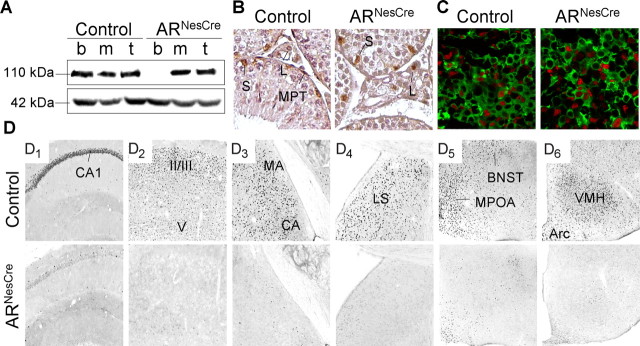
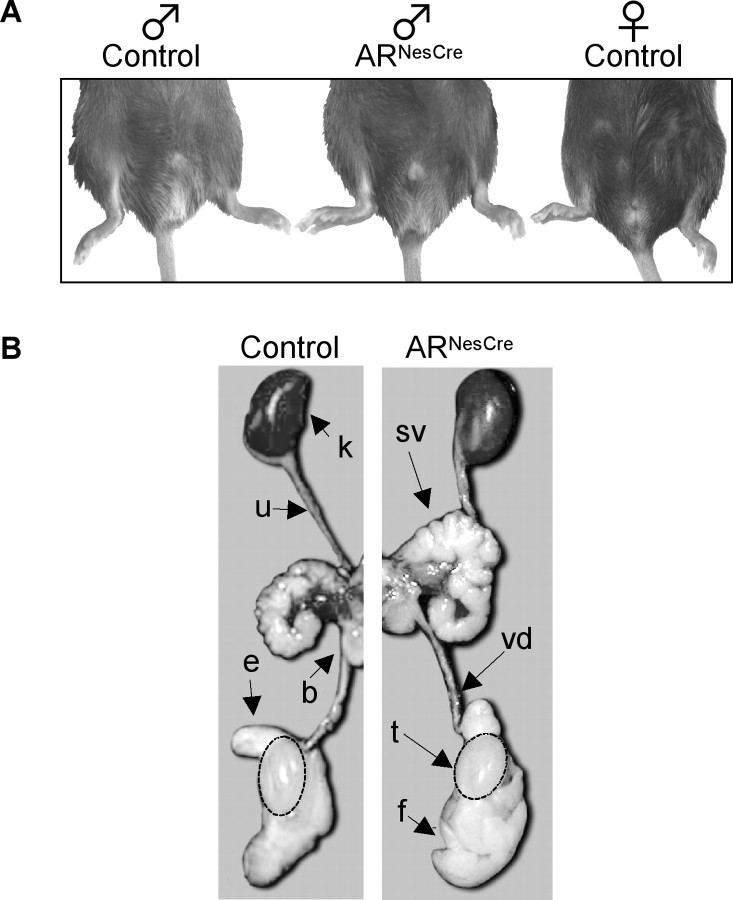
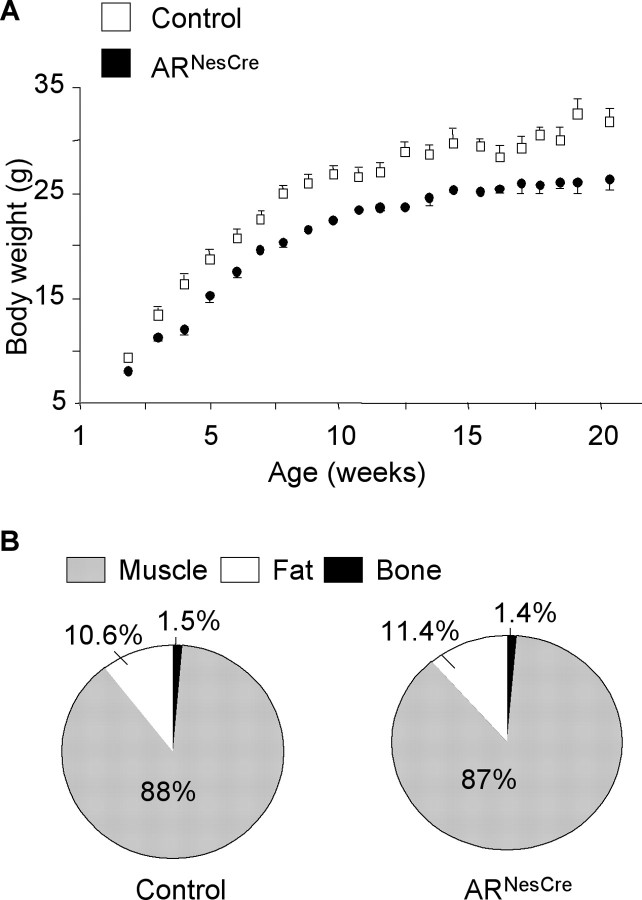
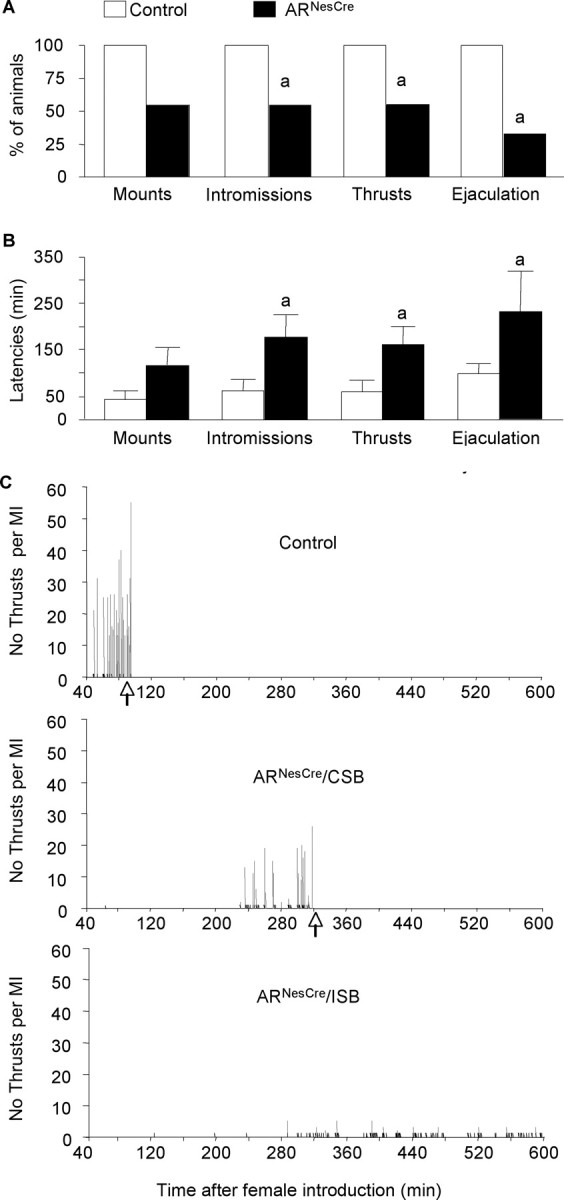
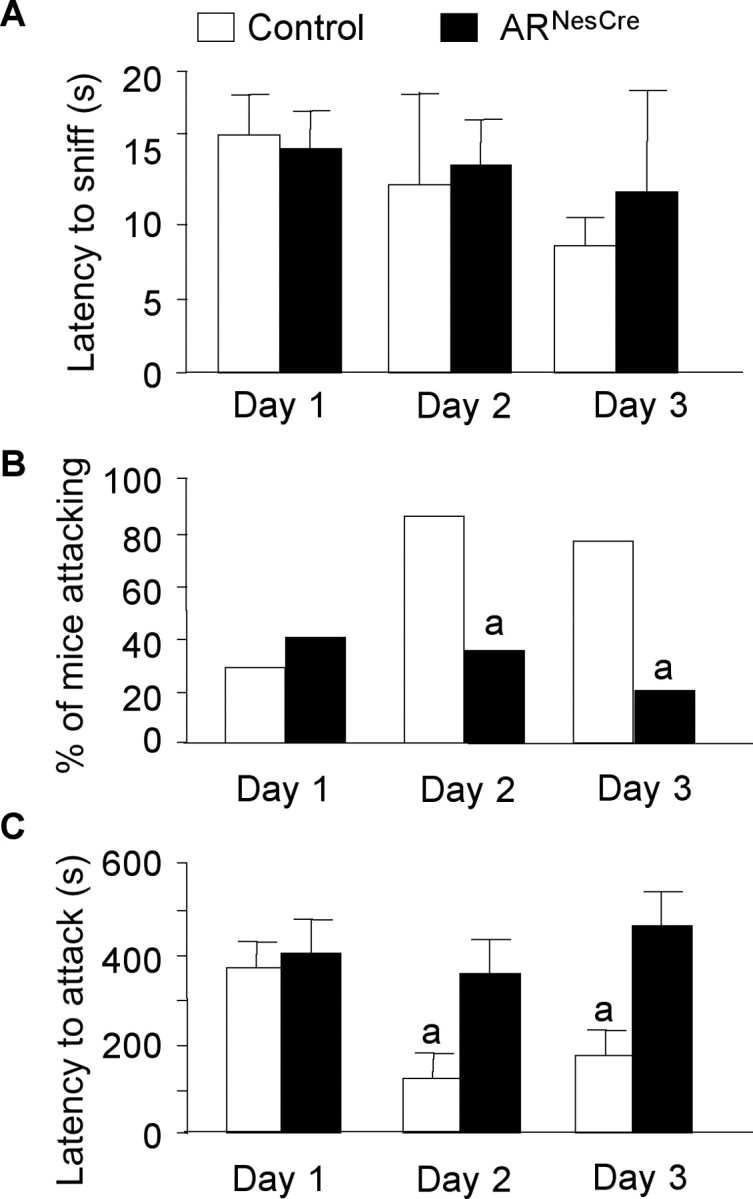
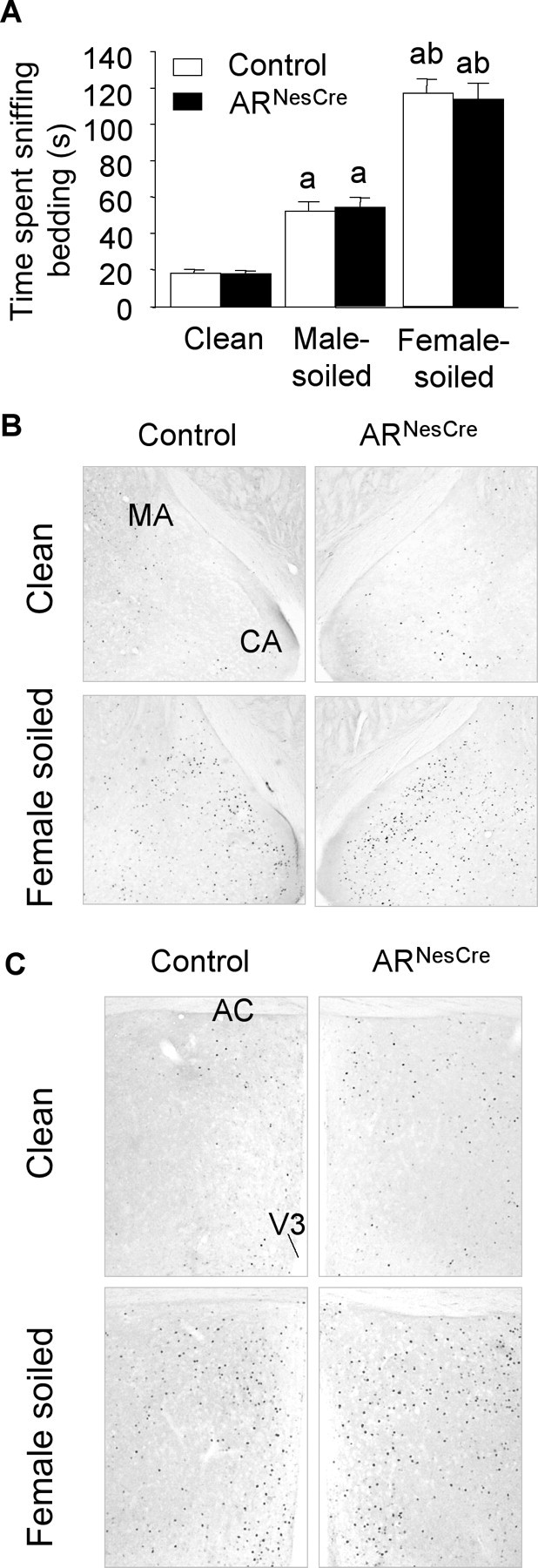
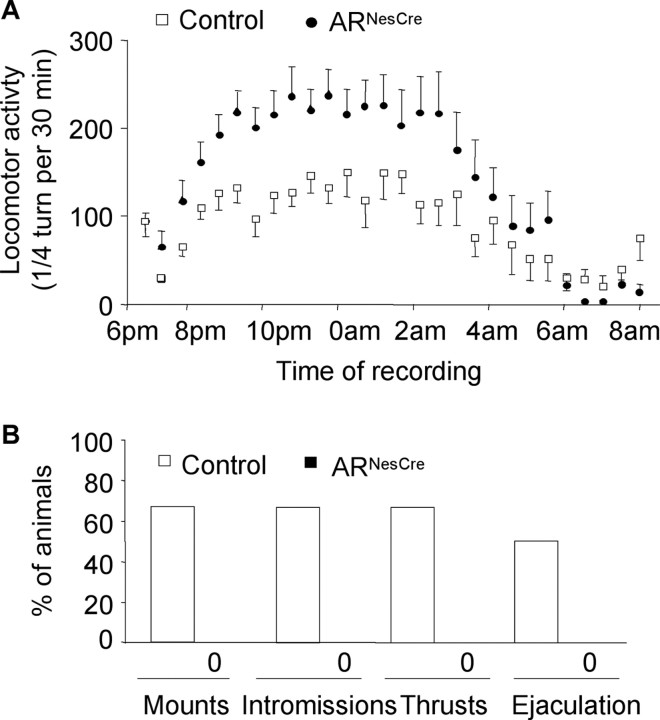
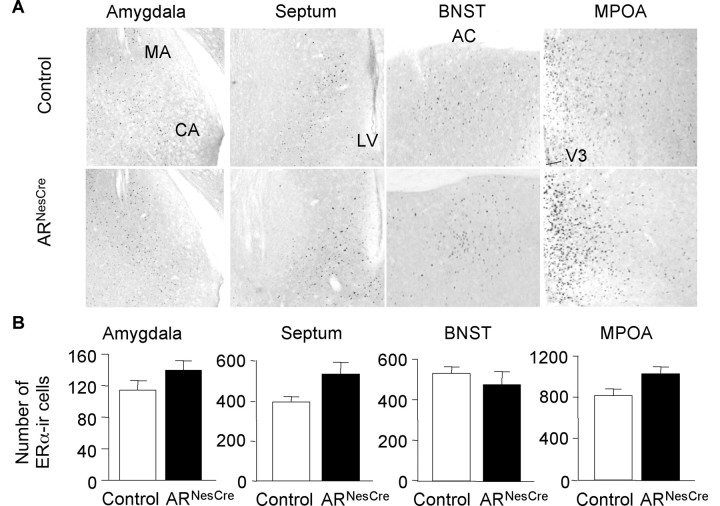
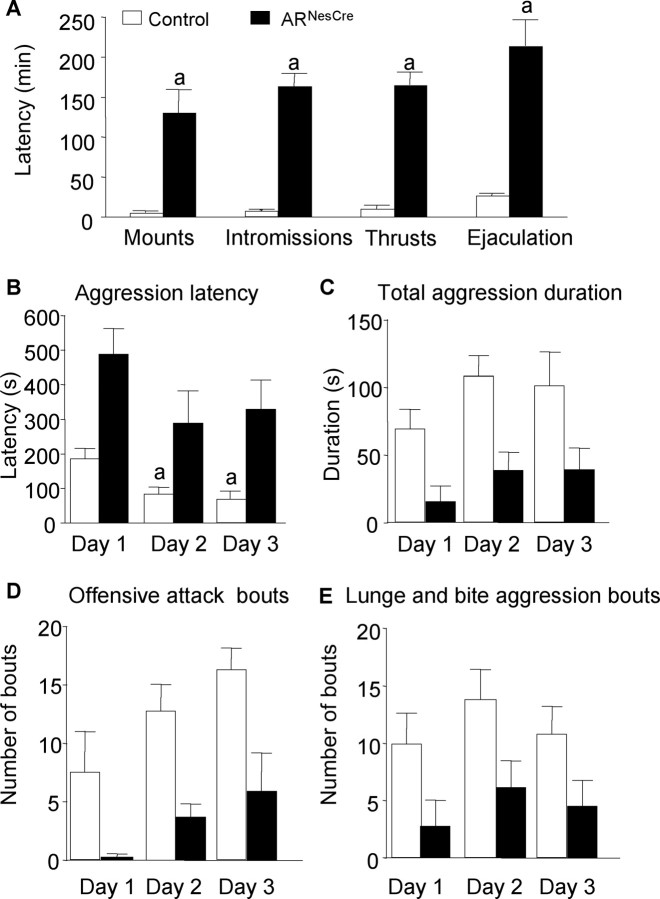
Testosterone (T) profoundly influences central sexual differentiation and functions. In the brain, T signals either directly through androgen receptor (AR) or indirectly through estrogen receptor (ER) following aromatization into E2 (17-beta-estradiol). As T, through AR, also controls peripheral male sexual differentiation, the relative contribution of central AR in T-mediated regulation of behavioral and neuroendocrine responses still remains unclear. To address this question, we generated, by using Cre-loxP technology, mice selectively lacking AR expression in the nervous system. The mutant male urogenital tract was normally developed, and mice were able to produce offspring. Nonetheless, sexual motivation and performance as well as aggressive behaviors were affected. Only a low percentage of males displayed a complete sexual behavior and offensive attacks. The latency to show masculine behaviors was increased and copulation length prolonged. Erectile activity during mating was also altered. These alterations occurred despite increased levels of T and its metabolites, and an unaffected number of ERalpha-immunoreactive cells. Olfactory preference and neuronal activation, mapped by Fos immunoreactivity, following exposure to estrus female-soiled bedding were also normal. At comparable T levels, greater differences in masculine behaviors were observed between gonadectomized control and mutant males. AR invalidation in the nervous system also disrupted the somatotropic axis since mutant males exhibited growth retardation and decreased serum levels of insulin-like growth factor I. Our findings show that central AR is required in T-induced regulation of male-typical behaviors and gonadotrope and somatotropic axes. This genetic model offers a unique opportunity in the understanding of AR's role in cerebral functions of T.
Figures
References
-
- Agmo A. Male rat sexual behavior. Brain Res Brain Res Protoc. 1997;1:203–209. - PubMed
-
- Arteaga-Silva M, Rodríguez-Dorantes M, Baig S, Morales-Montor J. Effects of castration and hormone replacement on male sexual behavior and pattern of expression in the brain of sex-steroid receptors in BALB/c AnN mice. Comp Biochem Physiol A Mol Integr Physiol. 2007;147:607–615. - PubMed
-
- Baker J, Hardy MP, Zhou J, Bondy C, Lupu F, Bellvé AR, Efstratiadis A. Effects of an Igf1 gene null mutation on mouse reproduction. Mol Endocrinol. 1996;10:903–918. - PubMed
-
- Bakker J, Honda S, Harada N, Balthazart J. Sexual partner preference requires a functional aromatase (cyp19) gene in male mice. Horm Behav. 2002;42:158–171. - PubMed
-
- Bodo C, Rissman EF. Androgen receptor is essential for sexual differentiation of responses to olfactory cues in mice. Eur J Neurosci. 2007;25:2182–2190. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials