

Excitation of histaminergic tuberomamillary neurons by thyrotropin-releasing hormone
- PMID: 19357273
- PMCID: PMC3198719
- DOI: 10.1523/JNEUROSCI.2976-08.2009
Excitation of histaminergic tuberomamillary neurons by thyrotropin-releasing hormone
Abstract
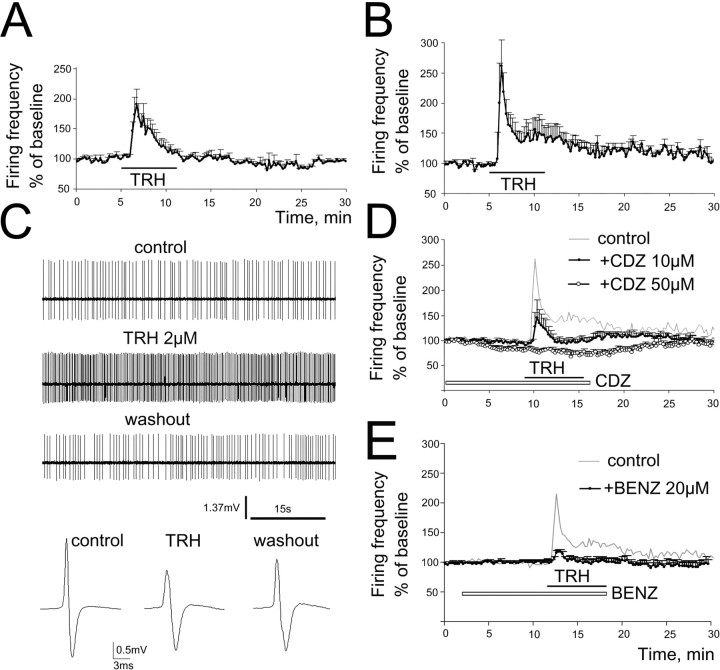
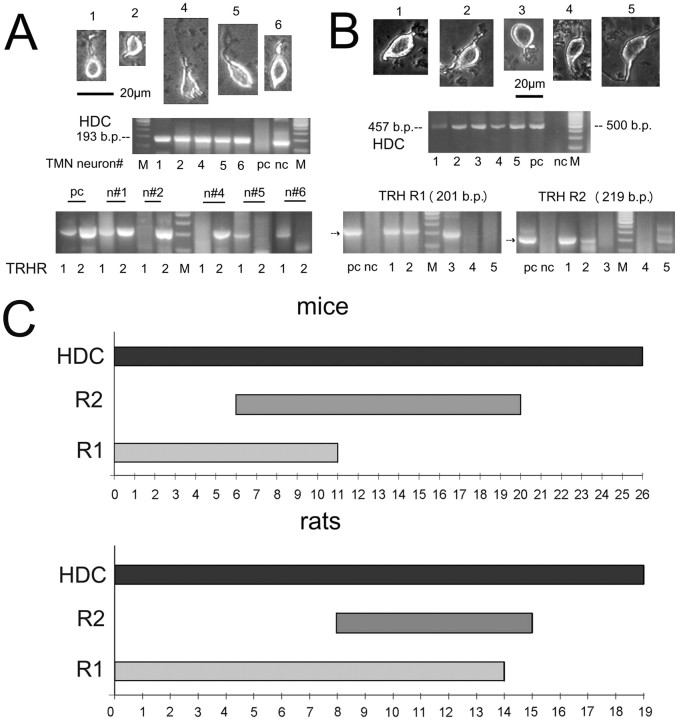
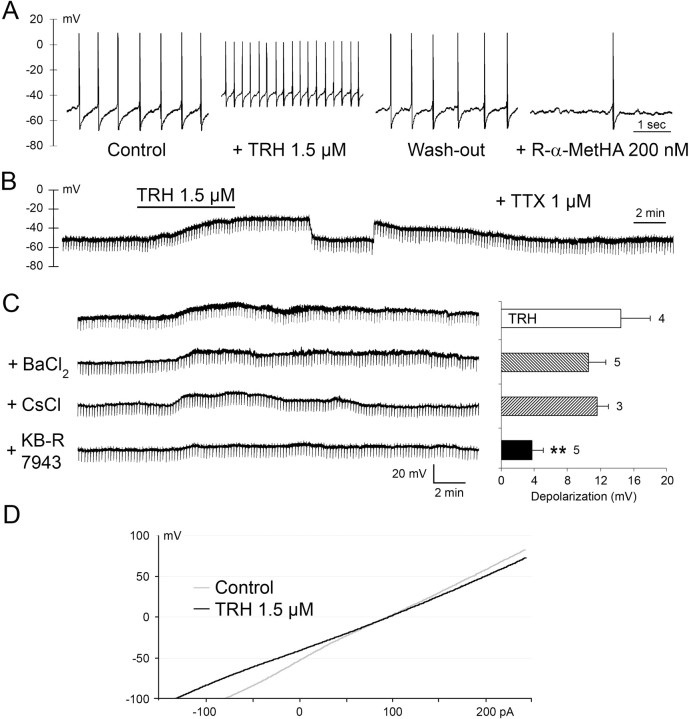
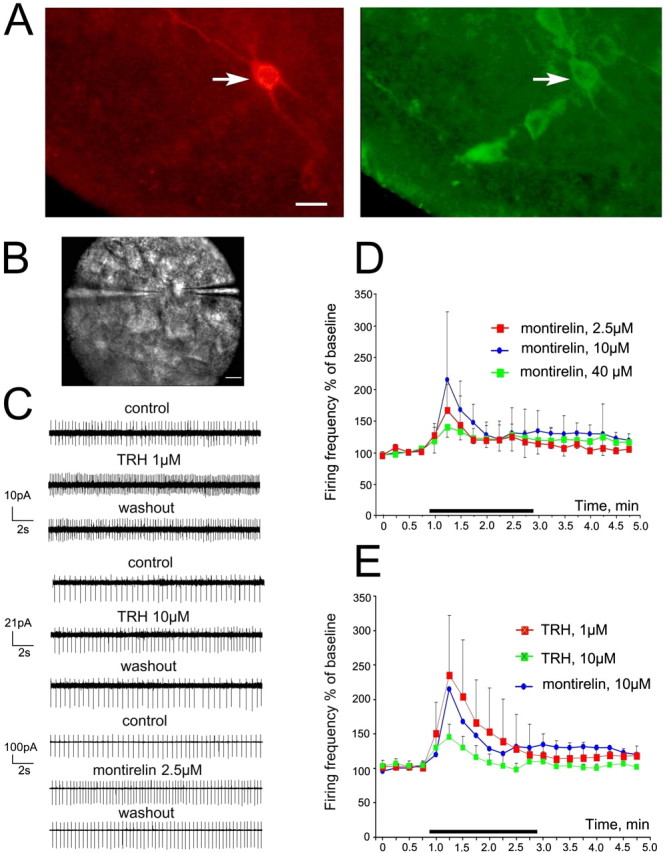
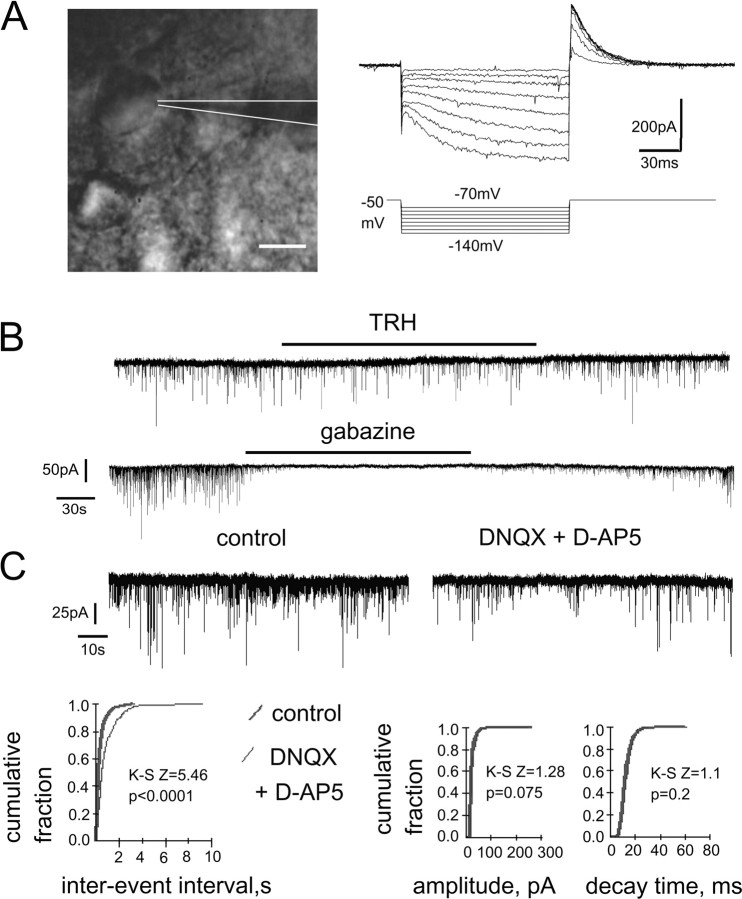
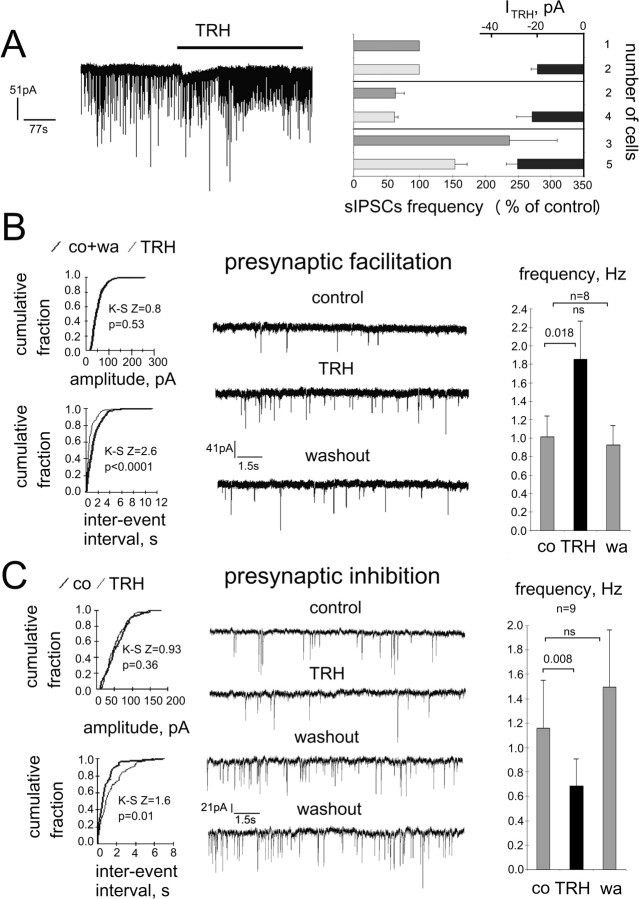
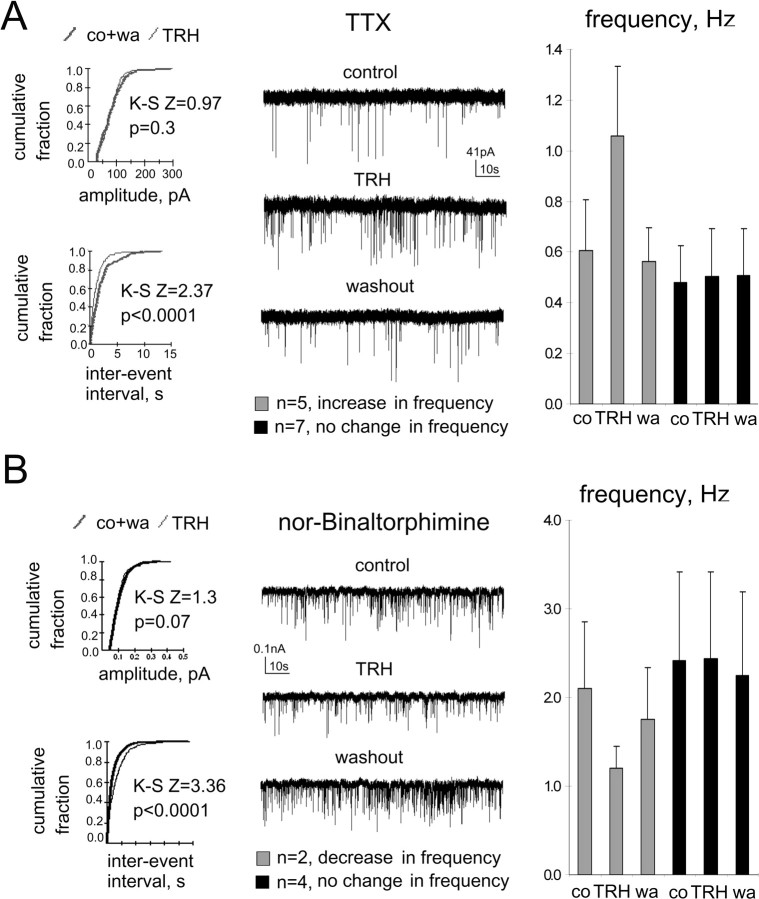
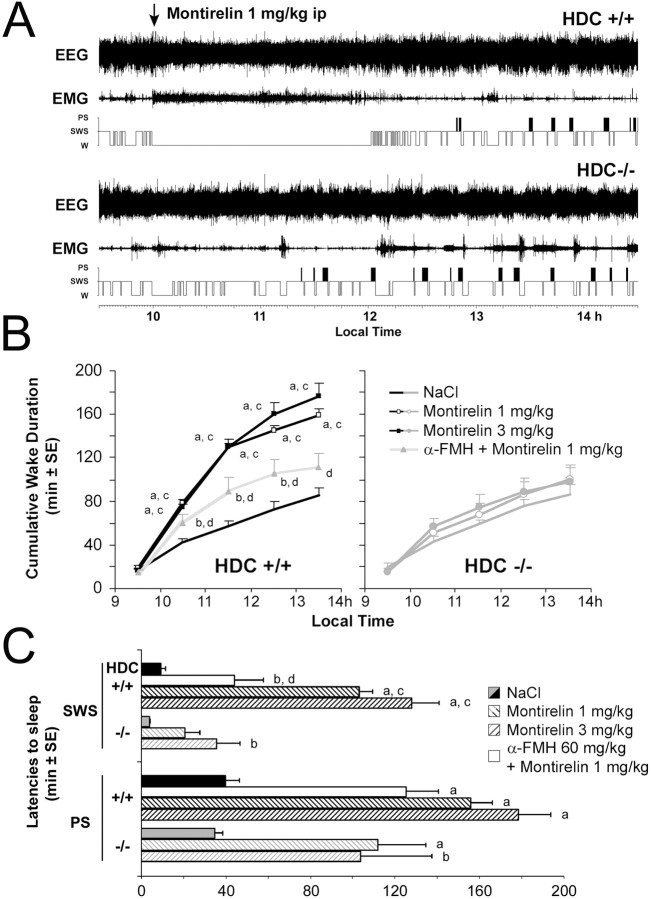
The histaminergic tuberomamillary nucleus (TMN) controls arousal and attention, and the firing of TMN neurons is state-dependent, active during waking, silent during sleep. Thyrotropin-releasing hormone (TRH) promotes arousal and combats sleepiness associated with narcolepsy. Single-cell reverse-transcription-PCR demonstrated variable expression of the two known TRH receptors in the majority of TMN neurons. TRH increased the firing rate of most (ca 70%) TMN neurons. This excitation was abolished by the TRH receptor antagonist chlordiazepoxide (CDZ; 50 mum). In the presence of tetrodotoxin (TTX), TRH depolarized TMN neurons without obvious change of their input resistance. This effect reversed at the potential typical for nonselective cation channels. The potassium channel blockers barium and cesium did not influence the TRH-induced depolarization. TRH effects were antagonized by inhibitors of the Na(+)/Ca(2+) exchanger, KB-R7943 and benzamil. The frequency of GABAergic spontaneous IPSCs was either increased (TTX-insensitive) or decreased [TTX-sensitive spontaneous IPSCs (sIPSCs)] by TRH, indicating a heterogeneous modulation of GABAergic inputs by TRH. Facilitation but not depression of sIPSC frequency by TRH was missing in the presence of the kappa-opioid receptor antagonist nor-binaltorphimine. Montirelin (TRH analog, 1 mg/kg, i.p.) induced waking in wild-type mice but not in histidine decarboxylase knock-out mice lacking histamine. Inhibition of histamine synthesis by (S)-alpha-fluoromethylhistidine blocked the arousal effect of montirelin in wild-type mice. We conclude that direct receptor-mediated excitation of rodent TMN neurons by TRH demands activation of nonselective cation channels as well as electrogenic Na(+)/Ca(2+) exchange. Our findings indicate a key role of the brain histamine system in TRH-induced arousal.
Figures
References
-
- Airaksinen MS, Alanen S, Szabat E, Visser TJ, Panula P. Multiple neurotransmitters in the tuberomammillary nucleus: comparison of rat, mouse, and guinea pig. J Comp Neurol. 1992;323:103–116. - PubMed
-
- Atzori M, Nistri A. Effects of thyrotropin-releasing hormone on GABAergic synaptic transmission of the rat hippocampus. Eur J Neurosci. 1996;8:1299–1305. - PubMed
-
- Bayliss DA, Viana F, Berger AJ. Mechanisms underlying excitatory effects of thyrotropin-releasing hormone on rat hypoglossal motoneurons in vitro. J Neurophysiol. 1992;68:1733–1745. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous