

CtIP-BRCA1 modulates the choice of DNA double-strand-break repair pathway throughout the cell cycle
- PMID: 19357644
- PMCID: PMC2857324
- DOI: 10.1038/nature07955
CtIP-BRCA1 modulates the choice of DNA double-strand-break repair pathway throughout the cell cycle
Abstract
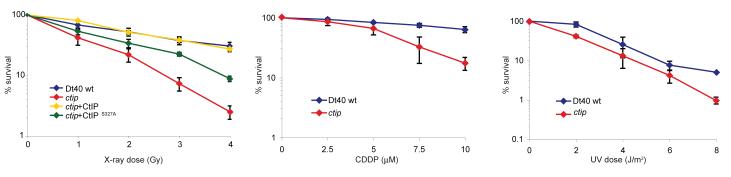
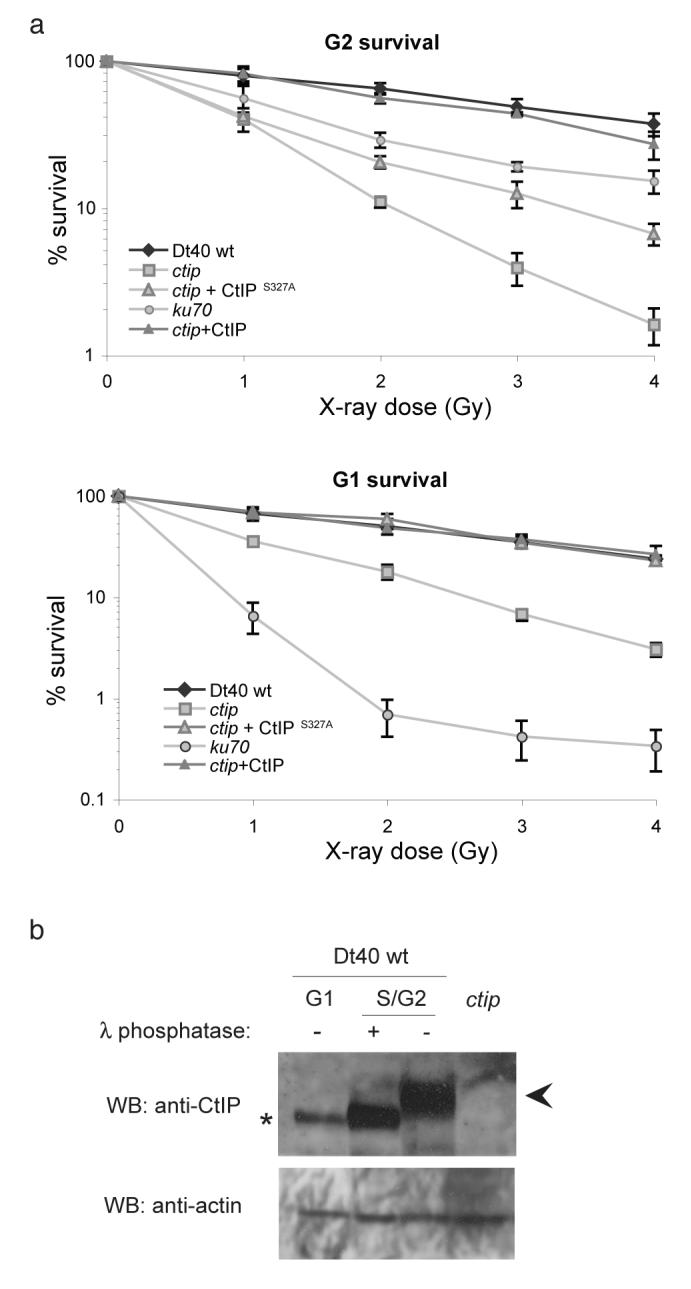
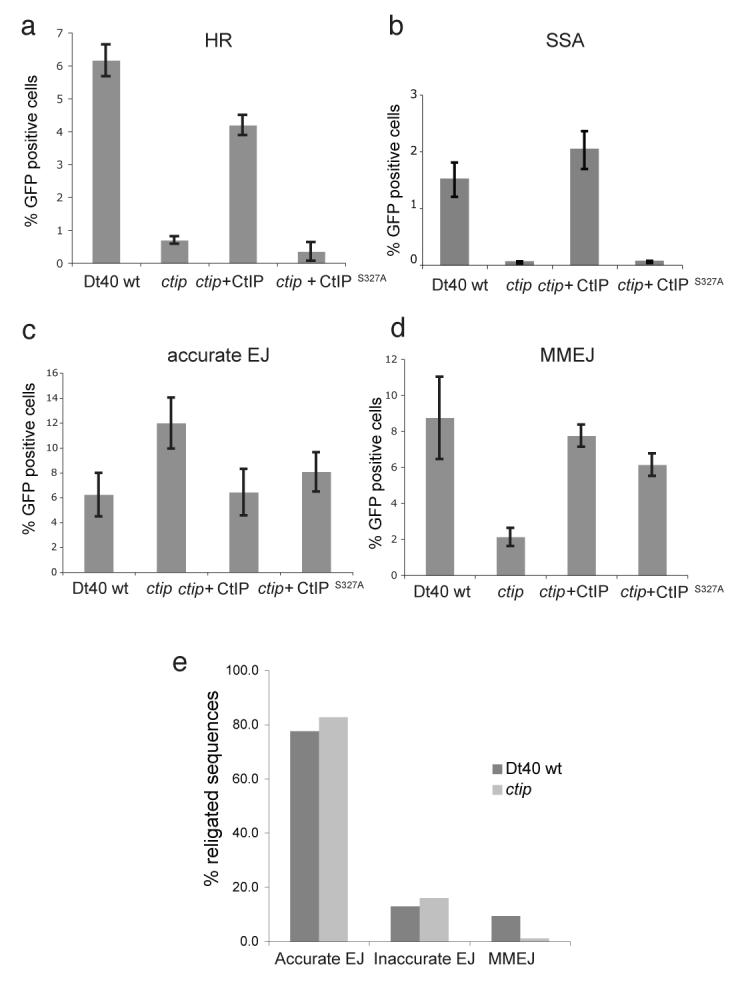
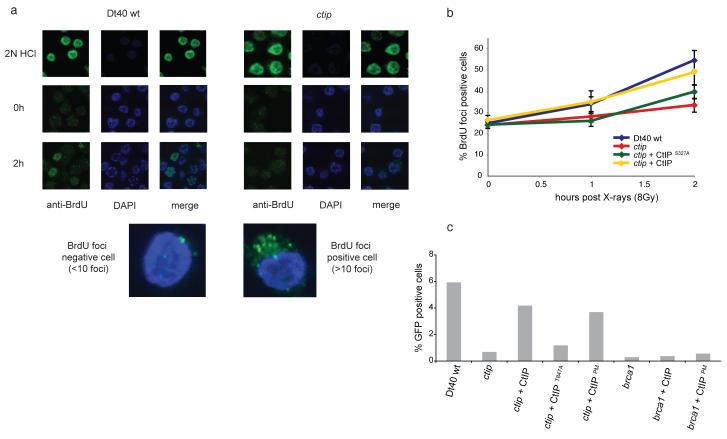
The repair of DNA double-strand breaks (DSBs) is tightly regulated during the cell cycle. In G1 phase, the absence of a sister chromatid means that repair of DSBs occurs through non-homologous end-joining or microhomology-mediated end-joining (MMEJ). These pathways often involve loss of DNA sequences at the break site and are therefore error-prone. In late S and G2 phases, even though DNA end-joining pathways remain functional, there is an increase in repair of DSBs by homologous recombination, which is mostly error-free. Consequently, the relative contribution of these different pathways to DSB repair in the cell cycle has a large influence on the maintenance of genetic integrity. It has remained unknown how DSBs are directed for repair by different, potentially competing, repair pathways. Here we identify a role for CtIP (also known as RBBP8) in this process in the avian B-cell line DT40. We establish that CtIP is required not only for repair of DSBs by homologous recombination in S/G2 phase but also for MMEJ in G1. The function of CtIP in homologous recombination, but not MMEJ, is dependent on the phosphorylation of serine residue 327 and recruitment of BRCA1. Cells expressing CtIP protein that cannot be phosphorylated at serine 327 are specifically defective in homologous recombination and have a decreased level of single-stranded DNA after DNA damage, whereas MMEJ remains unaffected. Our data support a model in which phosphorylation of serine 327 of CtIP as cells enter S phase and the recruitment of BRCA1 functions as a molecular switch to shift the balance of DSB repair from error-prone DNA end-joining to error-free homologous recombination.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
