

Thymosin beta4 mediated PKC activation is essential to initiate the embryonic coronary developmental program and epicardial progenitor cell activation in adult mice in vivo
- PMID: 19358334
- PMCID: PMC2768533
- DOI: 10.1016/j.yjmcc.2009.01.017
Thymosin beta4 mediated PKC activation is essential to initiate the embryonic coronary developmental program and epicardial progenitor cell activation in adult mice in vivo
Abstract
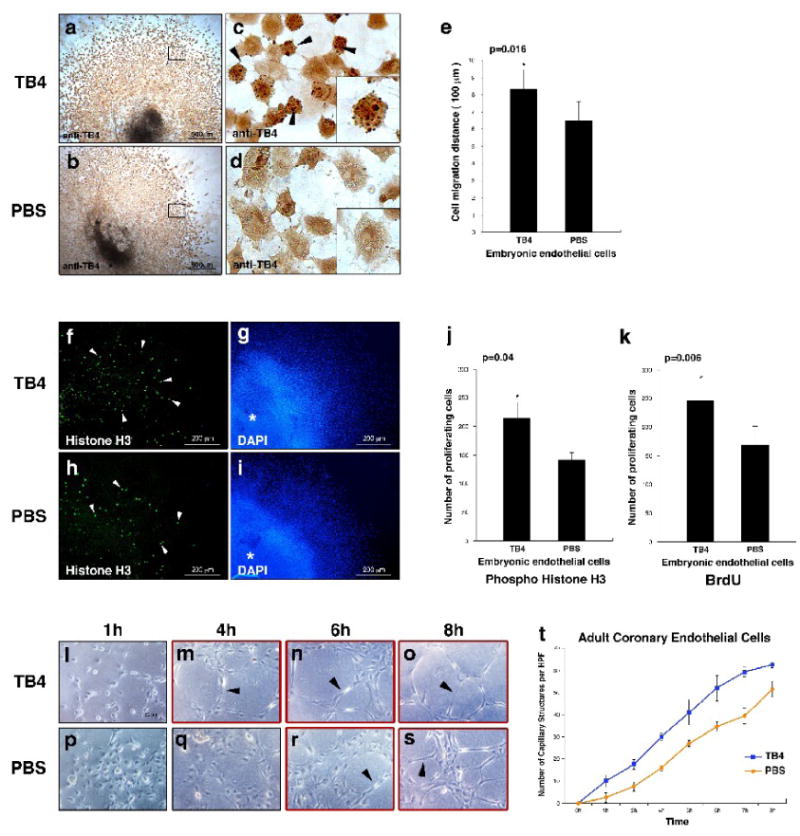
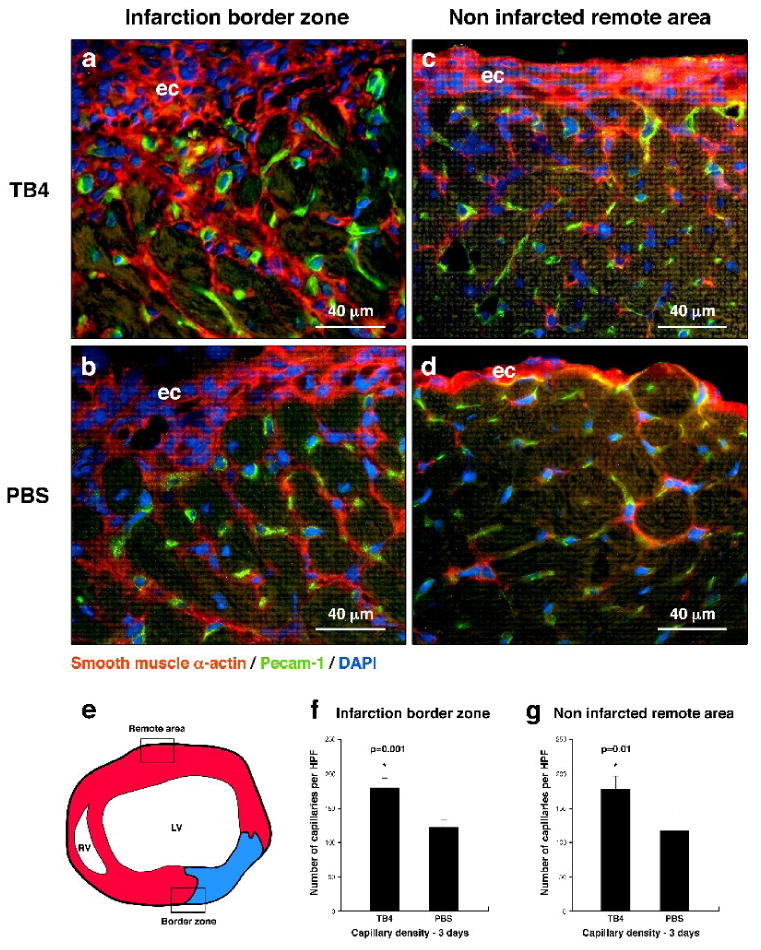
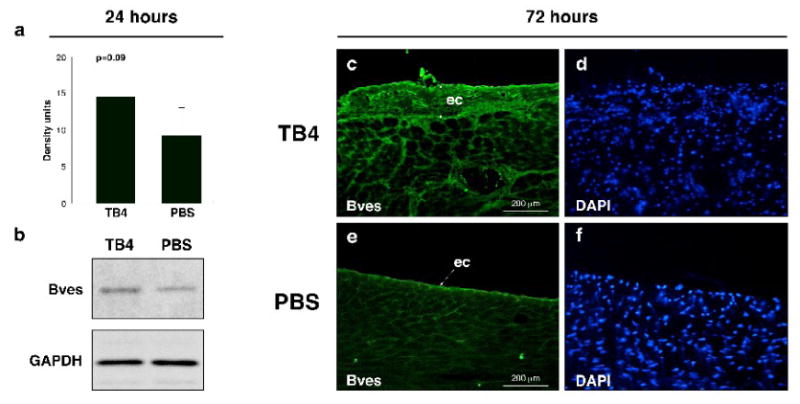
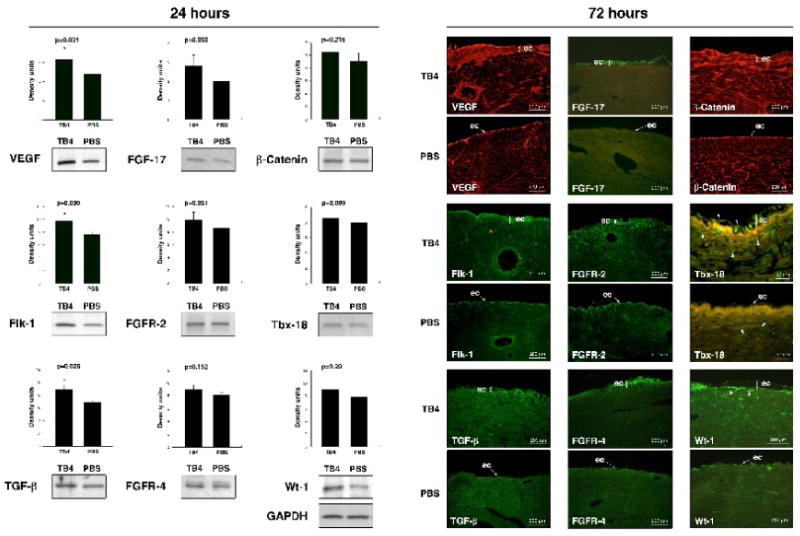
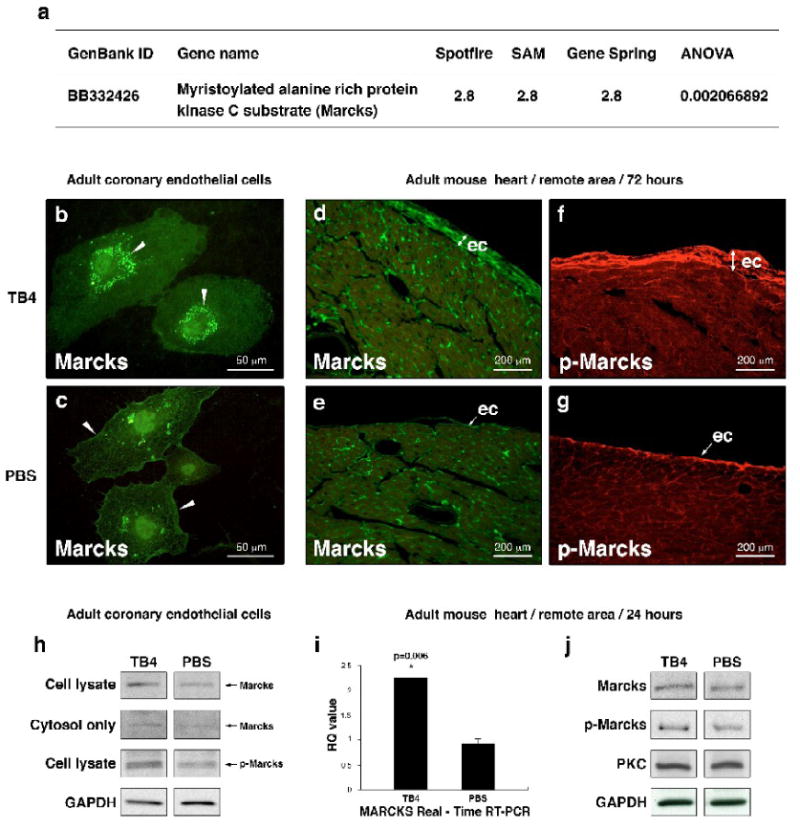
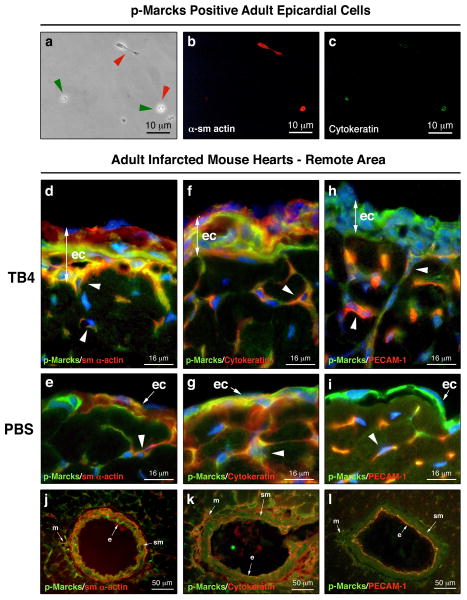
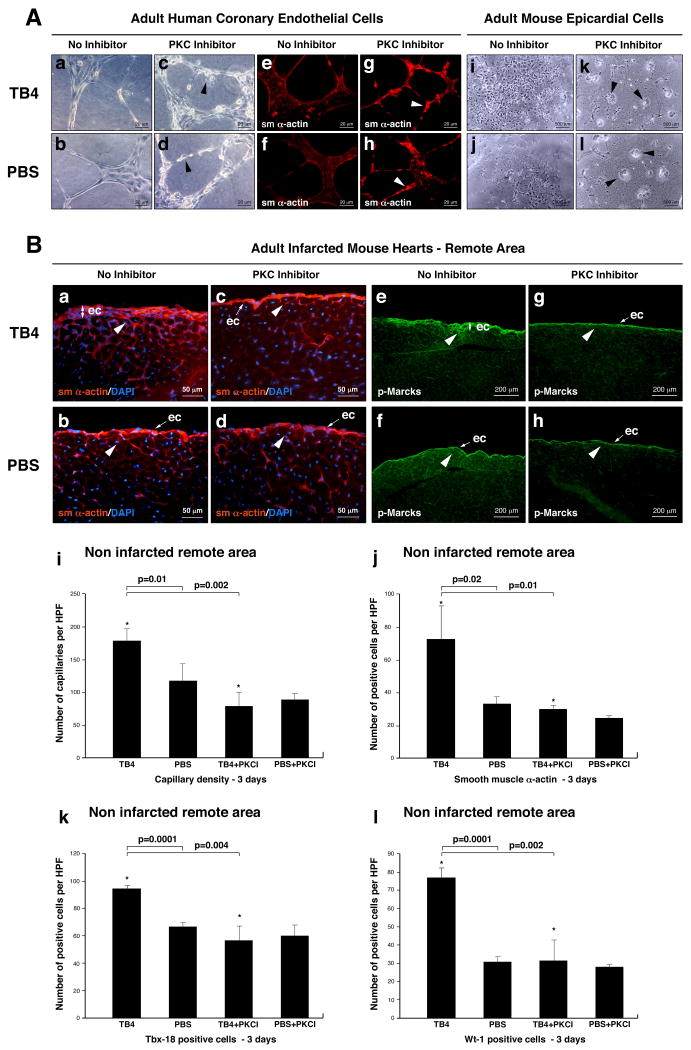
Hypoxic heart disease is a predominant cause of disability and death worldwide. Since adult mammalian hearts are incapable of regeneration after hypoxia, attempts to modify this deficiency are critical. As demonstrated in zebrafish, recall of the embryonic developmental program may be the key to success. Because thymosin beta4 (TB4) is beneficial for myocardial cell survival and essential for coronary development in embryos, we hypothesized that it reactivates the embryonic developmental program and initiates epicardial progenitor mobilization in adult mammals. We found that TB4 stimulates capillary-like tube formation of adult coronary endothelial cells and increases embryonic endothelial cell migration and proliferation in vitro. The increase of blood vessel/epicardial substance (Bves) expressing cells accompanied by elevated VEGF, Flk-1, TGF-beta, Fgfr-2, Fgfr-4, Fgf-17 and beta-Catenin expression and increase of Tbx-18 and Wt-1 positive myocardial progenitors suggested organ-wide recall of the embryonic program in the adult epicardium. TB4 also positively regulated the expression and phosphorylation of myristoylated alanine-rich C-kinase substrate (Marcks), a direct substrate and indicator of protein kinase C (PKC) activity in vitro and in vivo. PKC inhibition significantly reduced TB4 initiated epicardial thickening, capillary growth and the number of myocardial progenitors. Our results demonstrate that TB4 is the first known molecule capable of organ-wide activation of the embryonic coronary developmental program in the adult mammalian heart after systemic administration and that PKC plays a significant role in the process.
Figures
References
-
- Carmeliet P. Mechanisms of angiogenesis and arteriogenesis. Nat Med. 2000;6:389–395. - PubMed
-
- Tomanek RJ. Formation of the coronary vasculature during development. Angiogenesis. 2005;8:273–284. - PubMed
-
- Mu H, Ohashi R, Lin P, Yao Q, Chen C. Cellular and molecular mechanisms of coronary vessel development. Vasc Med. 2005;10:37–44. - PubMed
-
- Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, et al. Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell. 2003;114:763–776. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous