

A straight path to circular proteins
- PMID: 19359246
- PMCID: PMC2708896
- DOI: 10.1074/jbc.M901752200
A straight path to circular proteins
Abstract
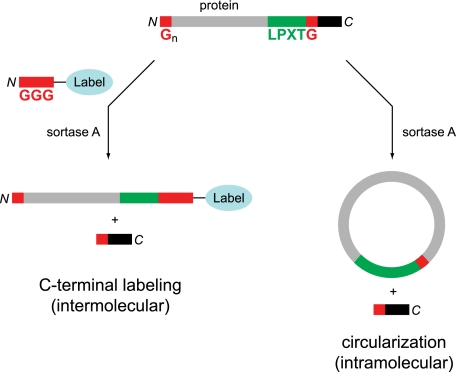
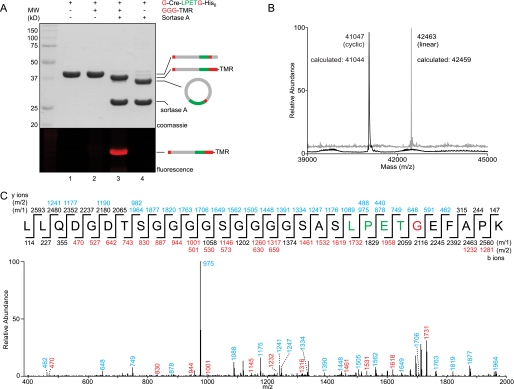
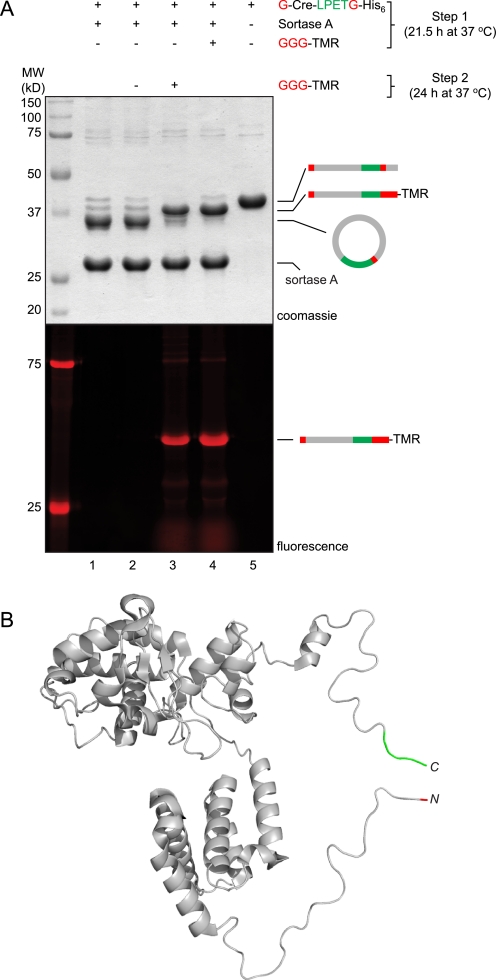
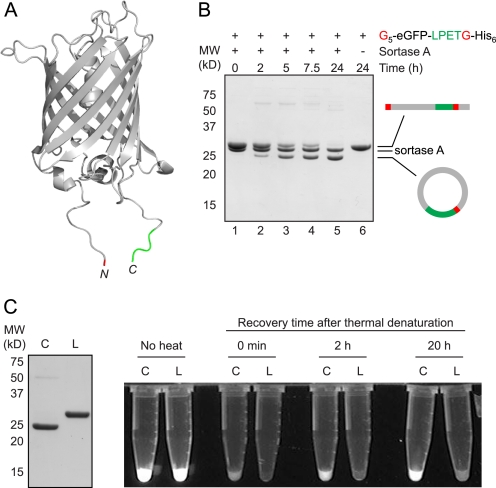
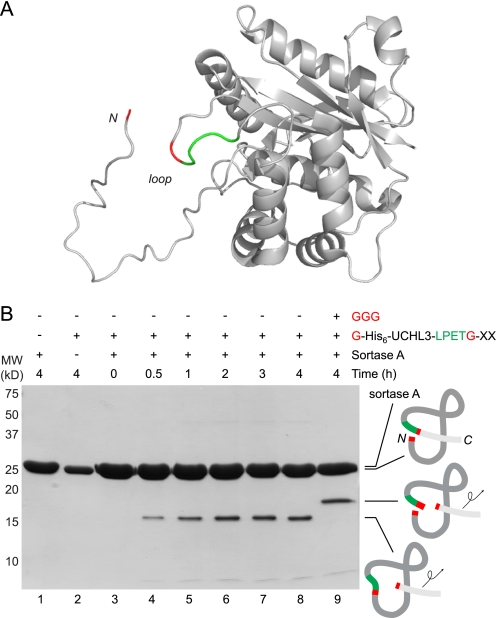
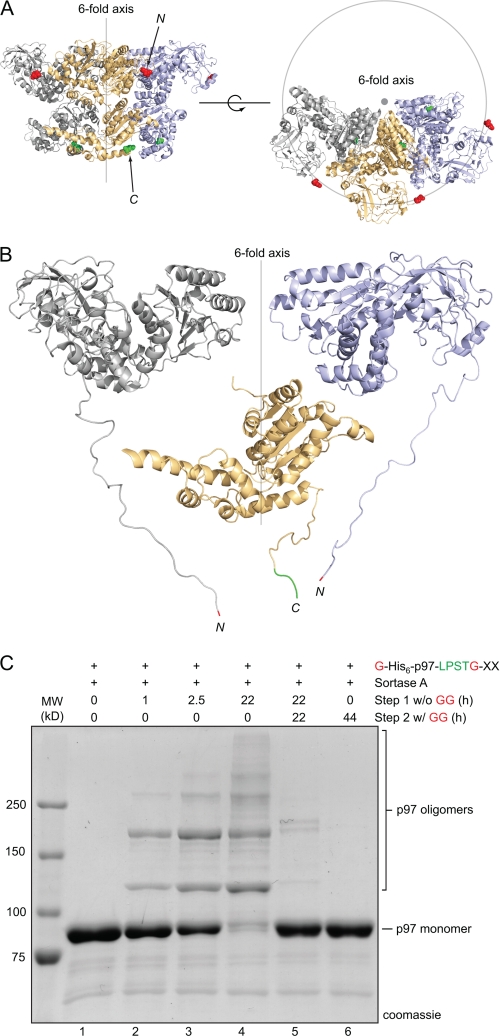
Folding and stability are parameters that control protein behavior. The possibility of conferring additional stability on proteins has implications for their use in vivo and for their structural analysis in the laboratory. Cyclic polypeptides ranging in size from 14 to 78 amino acids occur naturally and often show enhanced resistance toward denaturation and proteolysis when compared with their linear counterparts. Native chemical ligation and intein-based methods allow production of circular derivatives of larger proteins, resulting in improved stability and refolding properties. Here we show that circular proteins can be made reversibly with excellent efficiency by means of a sortase-catalyzed cyclization reaction, requiring only minimal modification of the protein to be circularized.
Figures
References
-
- Paterson G. K., Mitchell T. J. ( 2004) Trends Microbiol. 12, 89– 95 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
