

Ubx4 modulates cdc48 activity and influences degradation of misfolded proteins of the endoplasmic reticulum
- PMID: 19359248
- PMCID: PMC2713509
- DOI: 10.1074/jbc.M809282200
Ubx4 modulates cdc48 activity and influences degradation of misfolded proteins of the endoplasmic reticulum
Abstract
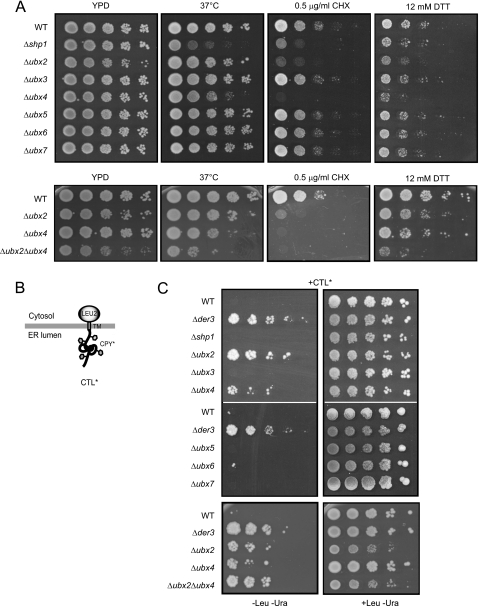
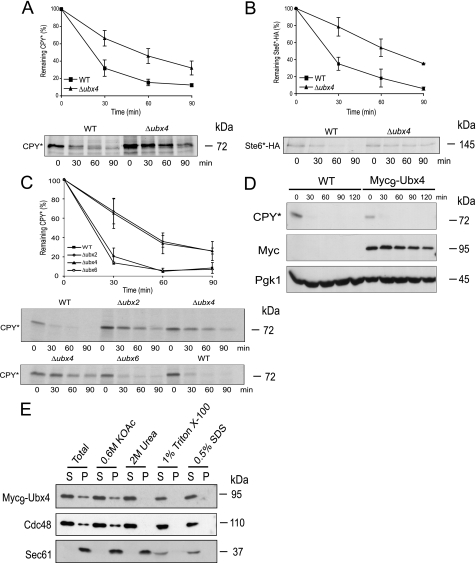
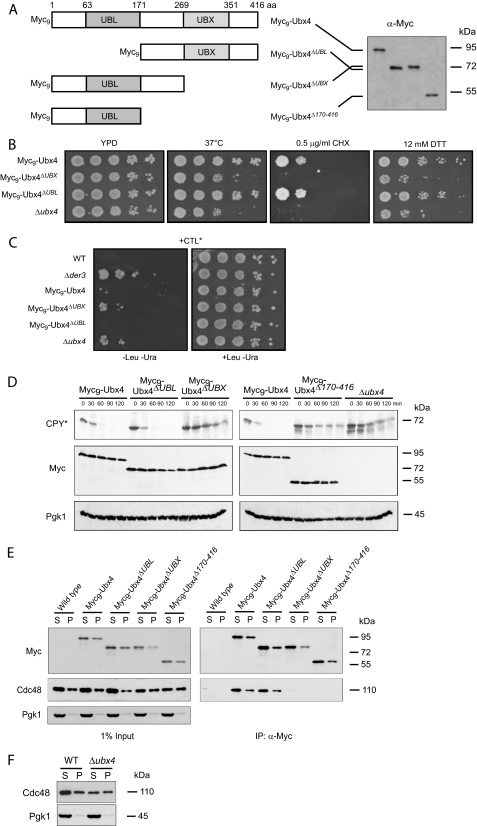
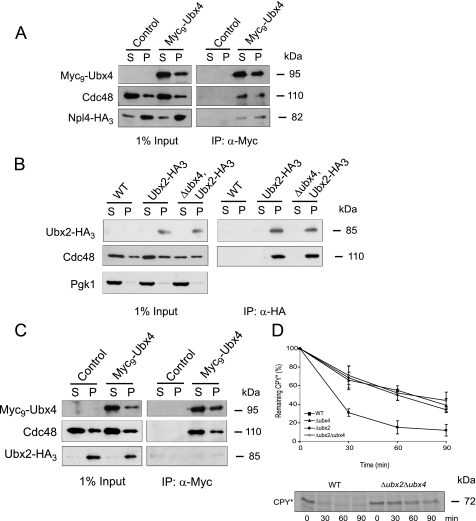
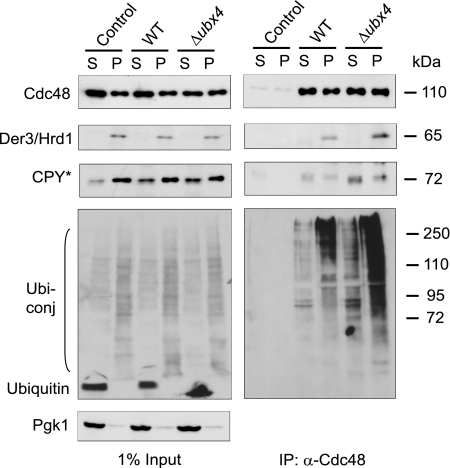
Misfolded proteins of the secretory pathway are recognized in the endoplasmic reticulum (ER), retrotranslocated into the cytoplasm, and degraded by the ubiquitin-proteasome system. Right after retrotranslocation and polyubiquitination, they are extracted from the cytosolic side of the ER membrane through a complex consisting of the AAA ATPase Cdc48 (p97 in mammals), Ufd1, and Npl4. This complex delivers misfolded proteins to the proteasome for final degradation. Extraction, delivery, and processing of ERAD (ER-associated degradation) substrates to the proteasome requires additional cofactors of Cdc48. Here we characterize the UBX domain containing protein Ubx4 (Cui1) as a crucial factor for the degradation of polyubiquitinated proteins via ERAD. Ubx4 modulates the Cdc48-Ufd1-Npl4 complex to guarantee its correct function. Mutant variants of Ubx4 lead to defective degradation of misfolded proteins and accumulation of polyubiquitinated proteins bound to Cdc48. We show the requirement of the UBX domain of Ubx4 for its function in ERAD. The observation that Ubx2 and Ubx4 are not found together in one complex with Cdc48 suggests several distinct steps in modulating the activity and localization of Cdc48 in ERAD.
Figures
References
-
- Hiller M. M., Finger A., Schweiger M., Wolf D. H. ( 1996) Science 273, 1725– 1728 - PubMed
-
- Ahner A., Brodsky J. L. ( 2004) Trends Cell Biol. 14, 474– 478 - PubMed
-
- Schäfer A., Kostova Z., Wolf D. H. ( 2008) in The Ubiquitin-Proteasome System and Disease, ( Mayer R. J., Ciechanover A., Rechsteiner M. eds) pp. 123– 143, Wiley-VCH, Weinheim, Germany
-
- Raasi S., Wolf D. H. ( 2007) Semin. Cell Dev. Biol. 18, 780– 791 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
