

doi: 10.1038/nrc2604.
Epub 2009 Apr 9.
The ups and downs of p53: understanding protein dynamics in single cells
Affiliations
- PMID: 19360021
- PMCID: PMC2892289
- DOI: 10.1038/nrc2604
Item in Clipboard
The ups and downs of p53: understanding protein dynamics in single cells
Nat Rev Cancer.
2009 May.
Abstract
Cells living in a complex environment must constantly detect, process and appropriately respond to changing signals. Therefore, all cellular information processing is dynamic in nature. As a consequence, understanding the process of signal transduction often requires detailed quantitative analysis of dynamic behaviours. Here, we focus on the oscillatory dynamics of the tumour suppressor protein p53 as a model for studying protein dynamics in single cells to better understand its regulation and function.
Figures
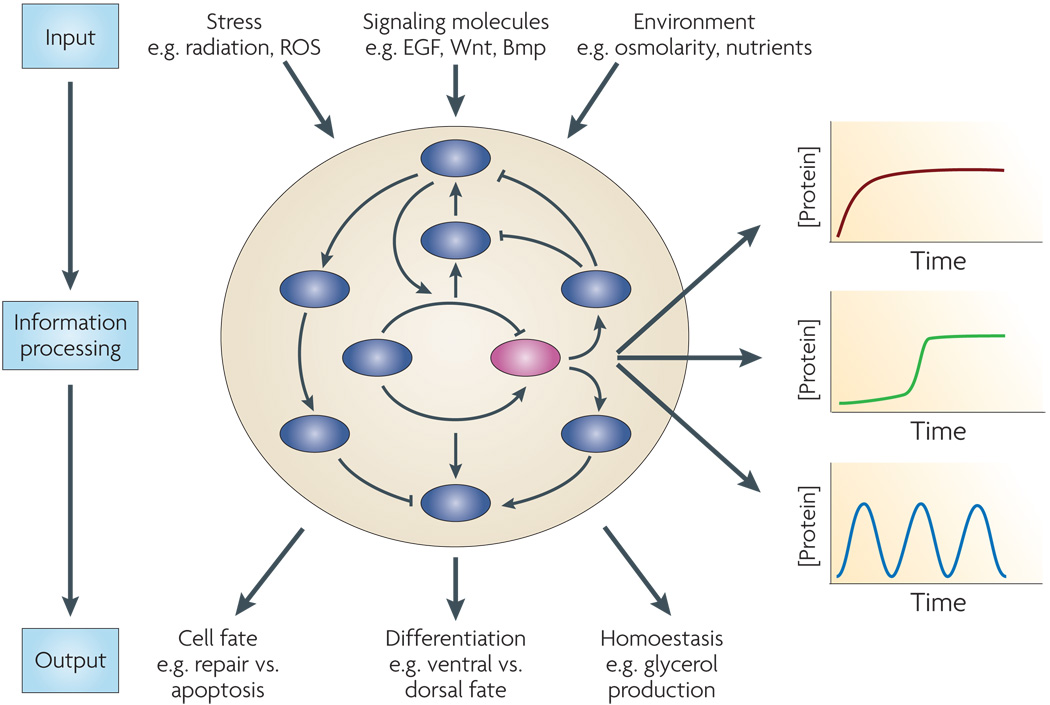
A complex protein network senses information about the intra- and extra-cellular environment (input), processes the information, and triggers a response (output). Currently, the information processing step is usually represented as a static drawing of binary (inhibitory or activating) arrows connecting different components of the network. An important aspect that is missing from such diagrams is the dynamical behavior of key members of the network. For example, the purple protein might show different dynamical behaviors in response to different inputs, in different cell types, or even between genetically identical cells. These dynamics can provide new insights about the specific interactions that are functional in each condition and the role of these interactions in triggering the right outcome.
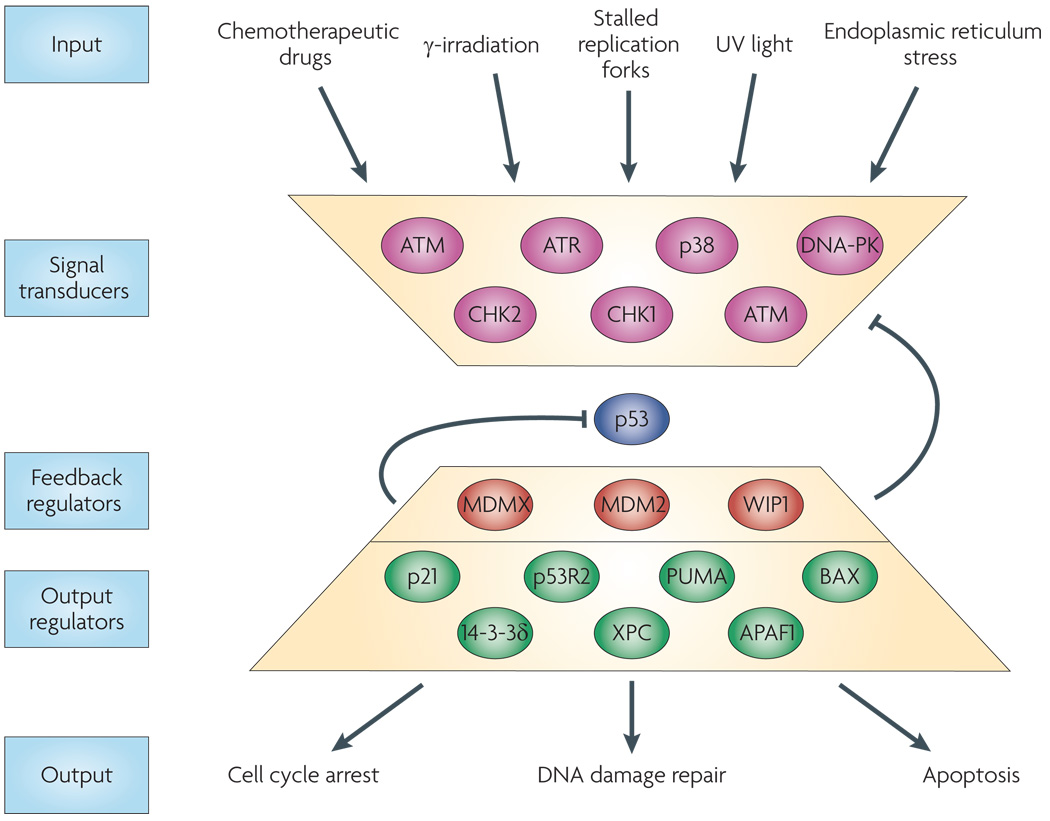
Stress signals (inputs) are detected and transduced to p53 through several kinases (signal trasducers). Upon activation, p53 upregulates the transcription of numerous genes. Some p53 targets act as “feedback regulators,” altering the activity of the kinases or the stability of p53. Other p53 targets are “output regulators” that trigger specific cellular outcomes, including cell cycle arrest and apoptosis.
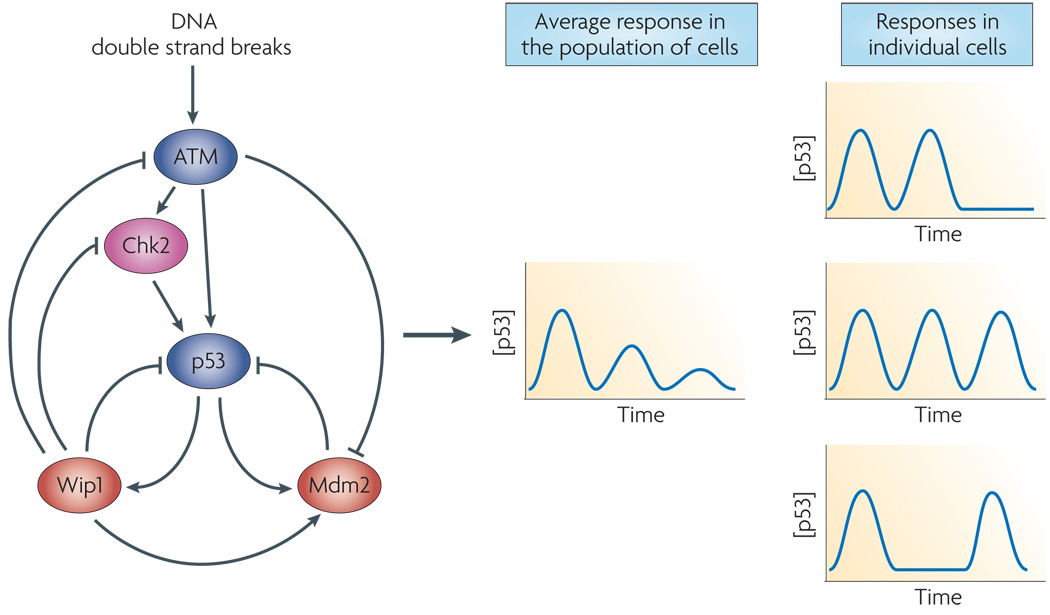
In response to DSBs ataxia telangiectasia mutated (ATM) kinase is activated, and activates checkpoint kinase 2 (Chk2), . Both of these kinases upregulate p53 by disrupting its interaction with one of its target genes, the E3 ubiquitin ligase Mdm2, , –. p53 also upregulates the transcription of the phosphatase Wip1 which negatively feeds back on the entire circuit by dephosphorylating ATM, Chk2, p53, and Mdm2. Solid lines represent protein-protein interactions, dashed lines represent transcriptional activation. Middle column; measurements averaged over populations of cells show damped oscillations of p53. Right column; single cell measurements show a series of undamped pulses with different cells showing different numbers of pulses.
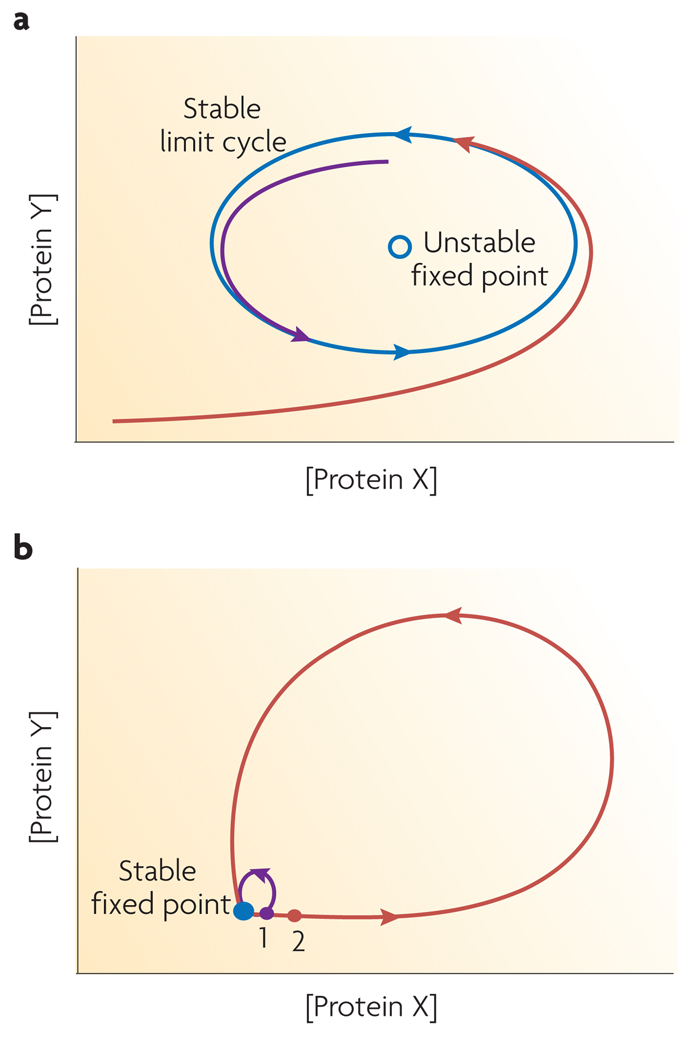
(a) The concentration of proteins composing an oscillator follow a cyclic path in phase space called a limit cycle (blue). The limit cycle is stable in that fluctuations of protein concentrations away from the limit cycle are suppressed. If the concentrations of the proteins in a system are initially different from values on the limit cycle, the system will relax away from unstable fixed points and will gradually approach the limit cycle. Two sample trajectories with different initial protein concentrations are shown in purple and red. (b) The concentration of proteins in a pulse generator will remain at a stable fixed point as long as the system is unperturbed. If the system is perturbed slightly, as indicated in point 1, it relaxes back to the stable point with relatively small changes in the protein concentrations (purple trajectory). However, if the system receives a large enough perturbation, such as to point 2, the concentrations of the proteins in the system change greatly, making a large excursion in phase space before relaxing back to the stable fixed point (red trajectory).
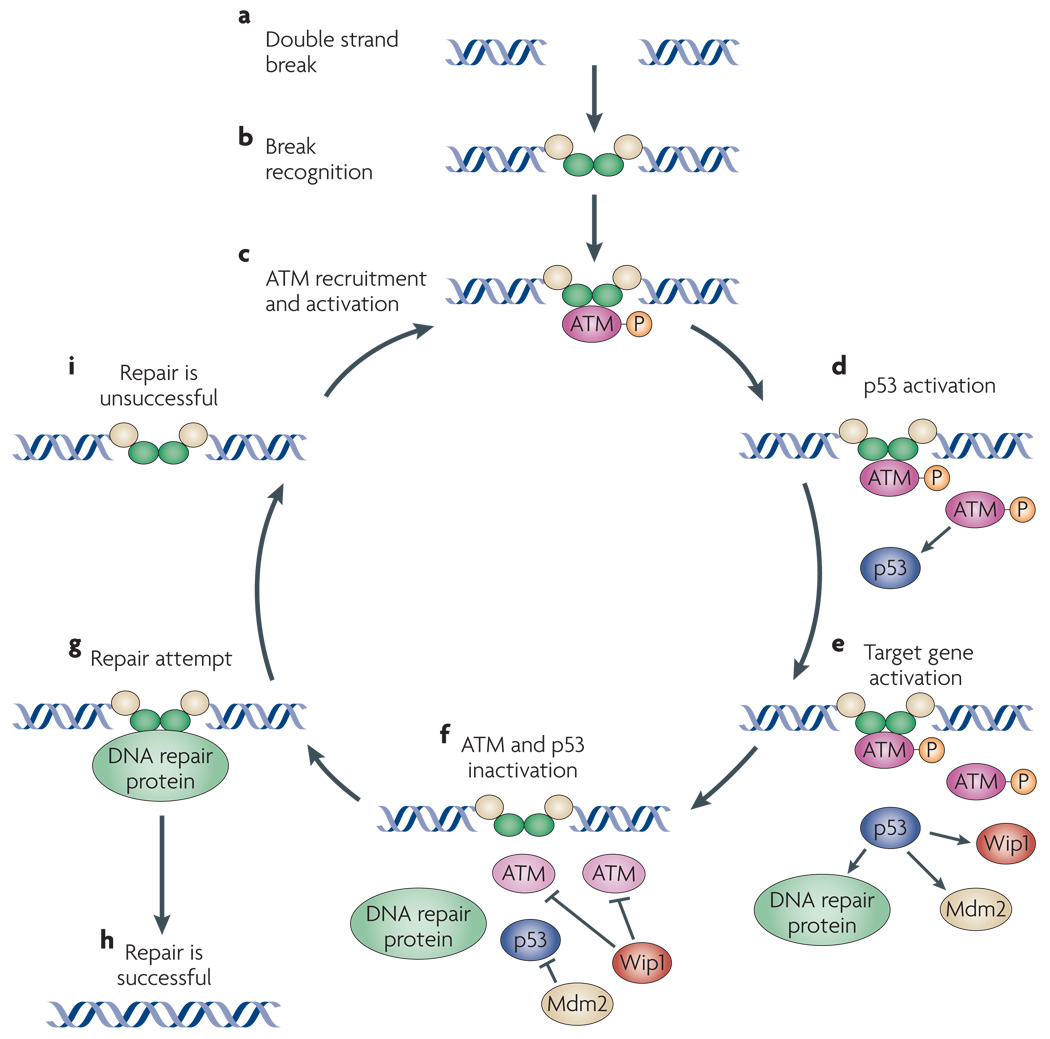
Upon occurrence of a DNA DSB (a), several proteins form a complex at the break site (b). ATM is recruited and activated by break recognition proteins (c). Once activated, ATM stabilizes p53 and triggers its accumulation. (d). p53 then transcriptionally upregulates the expression of several genes, including DNA repair genes and the inhibitor of ATM, Wip1 (e). Inhibition of ATM by Wip1 might lead to dissociation of ATM from the break (f), potentially allowing DNA repair proteins access to the DSB (g). If the DSB is repaired, ATM would not be recruited again and the signaling to p53 would be halted (h). If the DSB is not repaired (or if new breaks are detected), a new round of ATM recruitment would begin, resulting in a subsequent pulse of ATM and p53 (i).
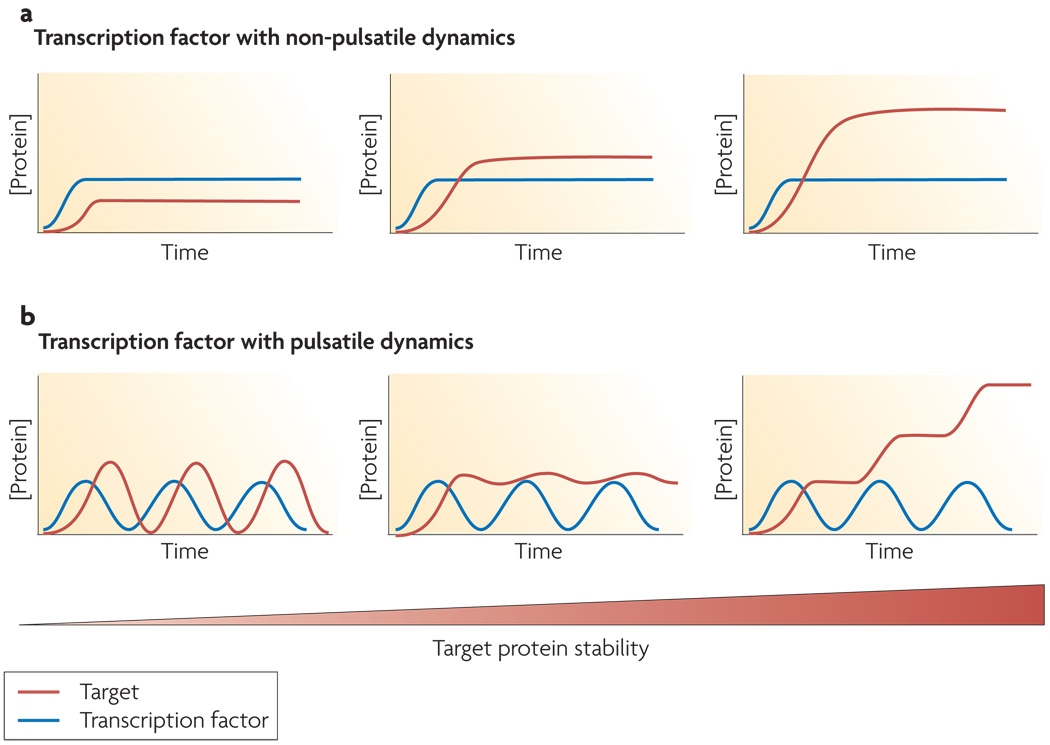
(a) If upon activation a transcription factor becomes elevated to a new concentration that remains constant in time, the level of its target genes will also remain constant in time with a steady state level that depends on the target protein’s stability (b) If a transcription factor undergoes pulses upon activation, the dynamics of its target genes can take on a wider range of qualitatively distinct behaviors. For example, targets can show pulsatile, steady, or step-like expression patterns depending on the stability of the target protein.
Similar articles
-
[Telomere maintenance by DNA damage response machinery].Seikagaku. 2010 Dec;82(12):1145-50. Seikagaku. 2010. PMID: 21348272 Review. Japanese. No abstract available.
-
[P53-independent apoptosis through a novel ATM/IKK-alpha/p73-mediated apoptotic pathway].Seikagaku. 2008 May;80(5):409-13. Seikagaku. 2008. PMID: 18575226 Review. Japanese. No abstract available.
-
[Activation of p53 protein after DNA damages].Tanpakushitsu Kakusan Koso. 2001 Jun;46(8 Suppl):1188-93. Tanpakushitsu Kakusan Koso. 2001. PMID: 11436309 Review. Japanese. No abstract available.
-
Cell fate decision mediated by p53 pulses.Proc Natl Acad Sci U S A. 2009 Jul 28;106(30):12245-50. doi: 10.1073/pnas.0813088106. Epub 2009 Jul 15. Proc Natl Acad Sci U S A. 2009. PMID: 19617533 Free PMC article.
-
The impact of a negligent G2/M checkpoint on genomic instability and cancer induction.Nat Rev Cancer. 2007 Nov;7(11):861-9. doi: 10.1038/nrc2248. Nat Rev Cancer. 2007. PMID: 17943134 Review.
Cited by
-
Clinicopathologic Correlations of E-cadherin and Prrx-1 Expression Loss in Hepatocellular Carcinoma.J Pathol Transl Med. 2016 Sep;50(5):327-36. doi: 10.4132/jptm.2016.06.22. Epub 2016 Aug 31. J Pathol Transl Med. 2016. PMID: 27580127 Free PMC article.
-
Misuse of the Michaelis-Menten rate law for protein interaction networks and its remedy.PLoS Comput Biol. 2020 Oct 22;16(10):e1008258. doi: 10.1371/journal.pcbi.1008258. eCollection 2020 Oct. PLoS Comput Biol. 2020. PMID: 33090989 Free PMC article. Review.
-
Toxicity testing in the 21 century: defining new risk assessment approaches based on perturbation of intracellular toxicity pathways.PLoS One. 2011;6(6):e20887. doi: 10.1371/journal.pone.0020887. Epub 2011 Jun 20. PLoS One. 2011. PMID: 21701582 Free PMC article. Review.
-
Transcription factor oscillations in neural stem cells: Implications for accurate control of gene expression.Neurogenesis (Austin). 2017 Feb 10;4(1):e1262934. doi: 10.1080/23262133.2016.1262934. eCollection 2017. Neurogenesis (Austin). 2017. PMID: 28321433 Free PMC article. Review.
-
Can case study approaches speed implementation of the NRC report: "Toxicity Testing in the 21st Century: A Vision and a Strategy?".ALTEX. 2011;28(3):175-82. doi: 10.14573/altex.2011.3.175. ALTEX. 2011. PMID: 21993955 Free PMC article.
References
-
- Cohen AA, et al. Dynamic proteomics of individual cancer cells in response to a drug. Science. 2008;322:1511–1516. - PubMed
-
- Hoffmann A, Levchenko A, Scott ML, Baltimore D. The IkappaB-NF-kappaB signaling module: temporal control and selective gene activation. Science. 2002;298:1241–1245. - PubMed
-
- Nelson DE, et al. Oscillations in NF-kappaB signaling control the dynamics of gene expression. Science. 2004;306:704–708. - PubMed
-
- Nelson DE, See V, Nelson G, White MR. Oscillations in transcription factor dynamics: a new way to control gene expression. Biochem Soc Trans. 2004;32:1090–1092. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
