

Electron microscopic evidence in support of alpha-solenoid models of proteasomal subunits Rpn1 and Rpn2
- PMID: 19361443
- PMCID: PMC2671960
- DOI: 10.1016/j.jmb.2009.01.039
Electron microscopic evidence in support of alpha-solenoid models of proteasomal subunits Rpn1 and Rpn2
Abstract
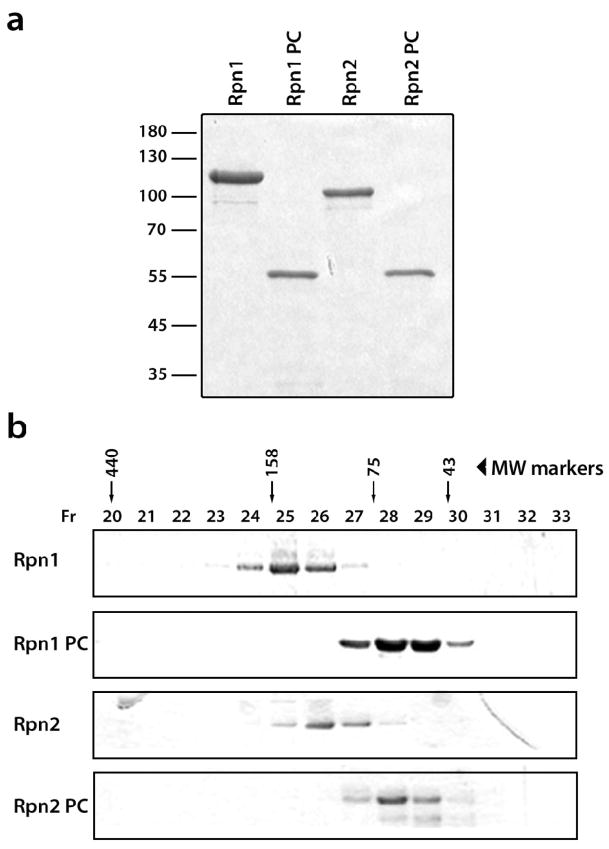
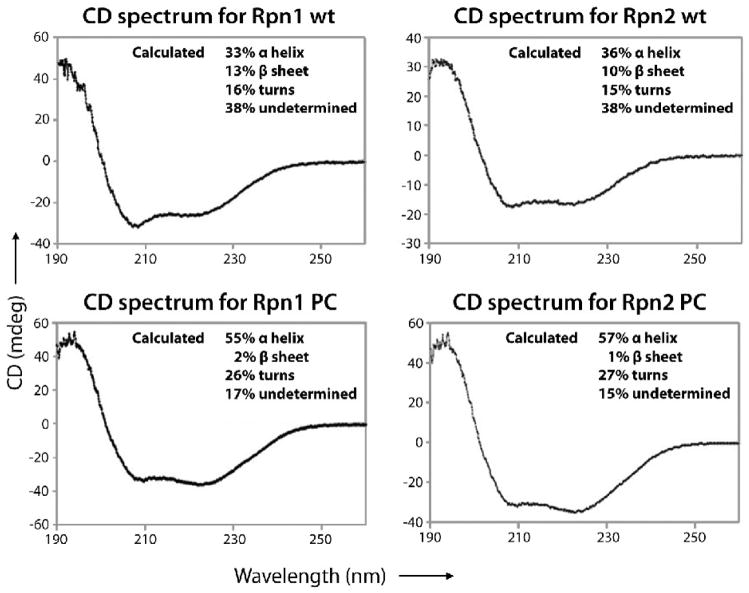
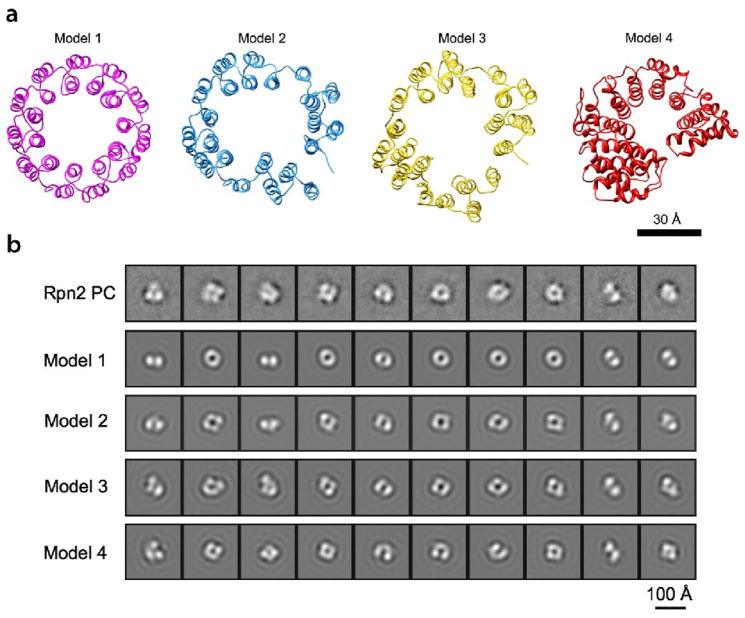
Rpn1 (109 kDa) and Rpn2 (104 kDa) are components of the 19S regulatory complex of the proteasome. The central portions of both proteins are predicted to have toroidal alpha-solenoid folds composed of 9-11 proteasome/cyclosome repeats, each approximately 40 residues long and containing two alpha-helices and turns [A. V. Kajava, J. Biol. Chem. 277, 49791-49798, 2002]. To evaluate this prediction, we examined the full-length yeast proteins and truncated versions thereof consisting only of the repeat-containing regions by gel filtration, CD spectroscopy, and negative-staining electron microscopy (EM). All four proteins are monomeric in solution and highly alpha-helical, particularly the truncated ones. The EM data were analyzed by image classification and averaging techniques. The preponderant projections, in each case, show near-annular molecules 6-7 nm in diameter. Comparison of the full-length with the truncated proteins showed molecules similar in size and shape, indicating that their terminal regions are flexible and thus smeared to invisibility in the averaged images. We tested the toroidal model further by calculating resolution-limited projections and comparing them with the EM images. The results support the alpha-solenoid model, except that they indicate that the repeats are organized not as symmetrical circular toroids but in less regular horseshoe-like structures.
Figures

References
-
- Zwickl P, Baumeister W, Steven A. Dis-assembly lines: the proteasome and related ATPase-assisted proteases. Curr Opin Struct Biol. 2000;10:242–50. - PubMed
-
- Goldberg AL. Protein degradation and protection against misfolded or damaged proteins. Nature. 2003;426:895–9. - PubMed
-
- Glickman MH, Rubin DM, Coux O, Wefes I, Pfeifer G, Cjeka Z, Baumeister W, Fried VA, Finley D. A subcomplex of the proteasome regulatory particle required for ubiquitin-conjugate degradation and related to the COP9-signalosome and eIF3. Cell. 1998;94:615–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
