

Levels of the ubiquitin ligase substrate adaptor MEL-26 are inversely correlated with MEI-1/katanin microtubule-severing activity during both meiosis and mitosis
- PMID: 19361490
- PMCID: PMC2720041
- DOI: 10.1016/j.ydbio.2009.04.004
Levels of the ubiquitin ligase substrate adaptor MEL-26 are inversely correlated with MEI-1/katanin microtubule-severing activity during both meiosis and mitosis
Abstract
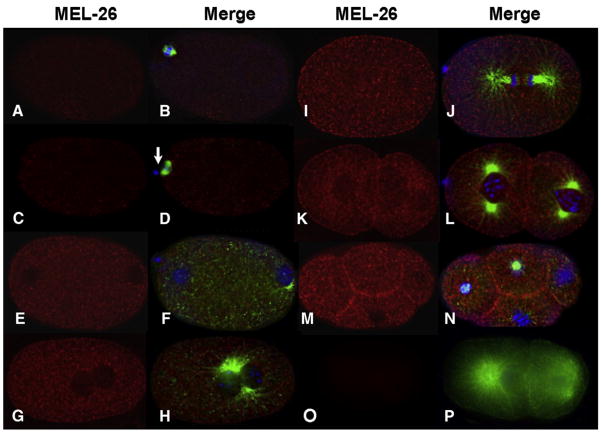
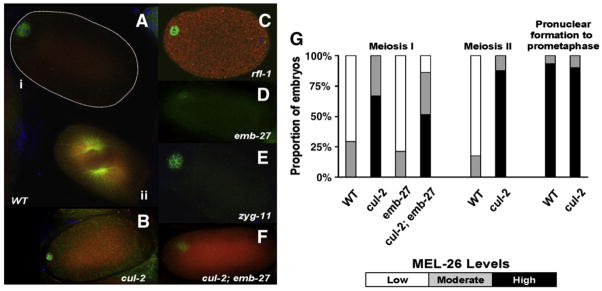
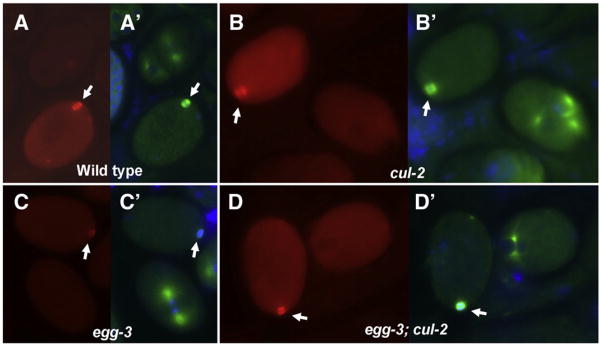
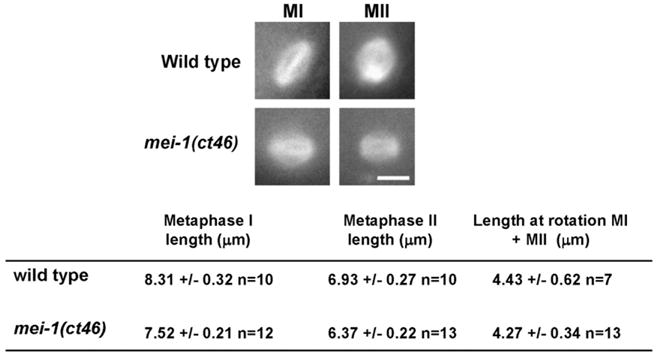
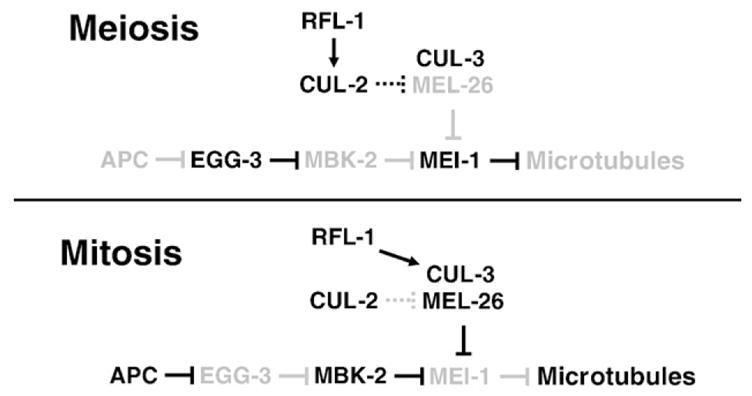
The MEI-1/MEI-2 microtubule-severing complex, katanin, is required for oocyte meiotic spindle formation and function in C. elegans, but the microtubule-severing activity must be quickly downregulated so that it does not interfere with formation of the first mitotic spindle. Post-meiotic MEI-1 inactivation is accomplished by two parallel protein degradation pathways, one of which requires MEL-26, the substrate-specific adaptor that recruits MEI-1 to a CUL-3 based ubiquitin ligase. Here we address the question of how MEL-26 mediated MEI-1 degradation is triggered only after the completion of MEI-1's meiotic function. We find that MEL-26 is present only at low levels until the completion of meiosis, after which protein levels increase substantially, likely increasing the post-meiotic degradation of MEI-1. During meiosis, MEL-26 levels are kept low by the action of another type of ubiquitin ligase, which contains CUL-2. However, we find that the low levels of meiotic MEL-26 have a subtle function, acting to moderate MEI-1 activity during meiosis. We also show that MEI-1 is the only essential target for MEL-26, and possibly for the E3 ubiquitin ligase CUL-3, but the upstream ubiquitin ligase activating enzyme RFL-1 has additional essential targets.
Figures
Similar articles
-
Regulation of the MEI-1/MEI-2 Microtubule-Severing Katanin Complex in Early Caenorhabditis elegans Development.G3 (Bethesda). 2016 Oct 13;6(10):3257-3268. doi: 10.1534/g3.116.031666. G3 (Bethesda). 2016. PMID: 27527792 Free PMC article.
-
The C. elegans anaphase promoting complex and MBK-2/DYRK kinase act redundantly with CUL-3/MEL-26 ubiquitin ligase to degrade MEI-1 microtubule-severing activity after meiosis.Dev Biol. 2007 Feb 15;302(2):438-47. doi: 10.1016/j.ydbio.2006.09.053. Epub 2006 Oct 5. Dev Biol. 2007. PMID: 17069791
-
Microtubule severing by the katanin complex is activated by PPFR-1-dependent MEI-1 dephosphorylation.J Cell Biol. 2013 Aug 5;202(3):431-9. doi: 10.1083/jcb.201304174. J Cell Biol. 2013. PMID: 23918937 Free PMC article.
-
Cellular Samurai: katanin and the severing of microtubules.J Cell Sci. 2000 Aug;113 ( Pt 16):2821-7. doi: 10.1242/jcs.113.16.2821. J Cell Sci. 2000. PMID: 10910766 Review.
-
Protein degradation: CUL-3 and BTB--partners in proteolysis.Curr Biol. 2004 Jan 20;14(2):R59-61. Curr Biol. 2004. PMID: 14738749 Review.
Cited by
-
The spindle assembly function of Caenorhabditis elegans katanin does not require microtubule-severing activity.Mol Biol Cell. 2011 May;22(9):1550-60. doi: 10.1091/mbc.E10-12-0951. Epub 2011 Mar 3. Mol Biol Cell. 2011. PMID: 21372175 Free PMC article.
-
UNC-89 (obscurin) binds to MEL-26, a BTB-domain protein, and affects the function of MEI-1 (katanin) in striated muscle of Caenorhabditis elegans.Mol Biol Cell. 2012 Jul;23(14):2623-34. doi: 10.1091/mbc.E12-01-0055. Epub 2012 May 23. Mol Biol Cell. 2012. PMID: 22621901 Free PMC article.
-
Regulation of the MEI-1/MEI-2 Microtubule-Severing Katanin Complex in Early Caenorhabditis elegans Development.G3 (Bethesda). 2016 Oct 13;6(10):3257-3268. doi: 10.1534/g3.116.031666. G3 (Bethesda). 2016. PMID: 27527792 Free PMC article.
-
The Mammalian Family of Katanin Microtubule-Severing Enzymes.Front Cell Dev Biol. 2021 Aug 3;9:692040. doi: 10.3389/fcell.2021.692040. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34414183 Free PMC article. Review.
-
Control of asymmetric cell division in early C. elegans embryogenesis: teaming-up translational repression and protein degradation.BMB Rep. 2010 Feb;43(2):69-78. doi: 10.5483/bmbrep.2010.43.2.069. BMB Rep. 2010. PMID: 20193124 Free PMC article. Review.
References
-
- Albertson DG. Formation of the first cleavage spindle in nematode embryos. Dev Biol. 1984;101:61–72. - PubMed
-
- Albertson DG, Thomson JN. Segregation of holocentric chromosomes at meiosis in the nematode, Caenorhabditis elegans. Chromos Res. 1993;1:15–26. - PubMed
-
- Bashir T, Dorrello NV, Amador V, Guardavaccaro D, Pagano M. Control of the SCF(Skp2-Cks1) ubiquitin ligase by the APC/C(Cdh1) ubiquitin ligase. Nature. 2004;428:190–193. - PubMed
-
- Bowerman B, Kurz T. Degrade to create: developmental requirements for ubiquitin-mediated proteolysis during early C. elegans embryogenesis. Development. 2006;133:773–784. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
