

Sonic hedgehog maintains proliferation in secondary heart field progenitors and is required for normal arterial pole formation
- PMID: 19361493
- PMCID: PMC2810612
- DOI: 10.1016/j.ydbio.2009.03.028
Sonic hedgehog maintains proliferation in secondary heart field progenitors and is required for normal arterial pole formation
Abstract
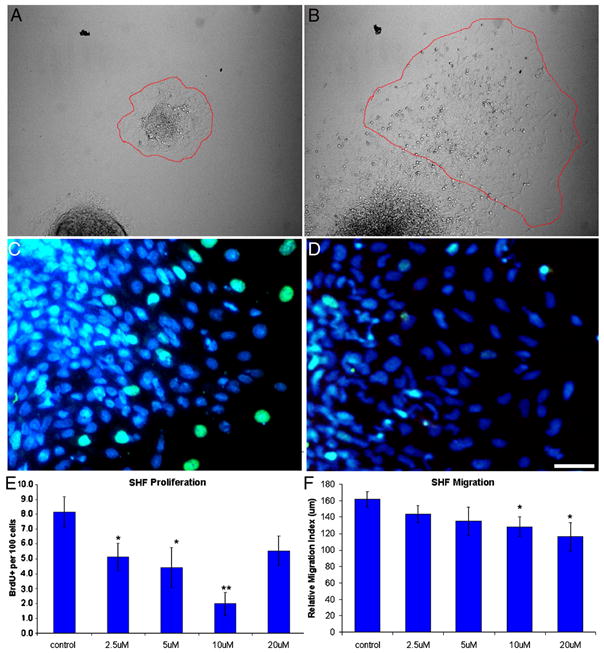
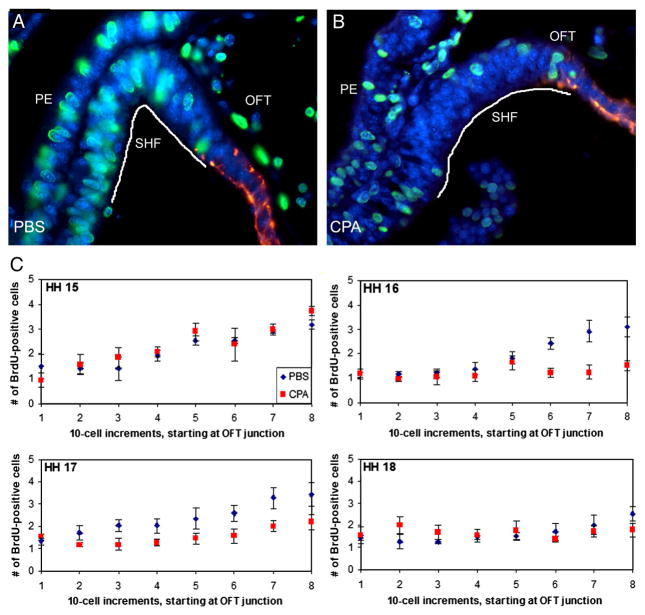
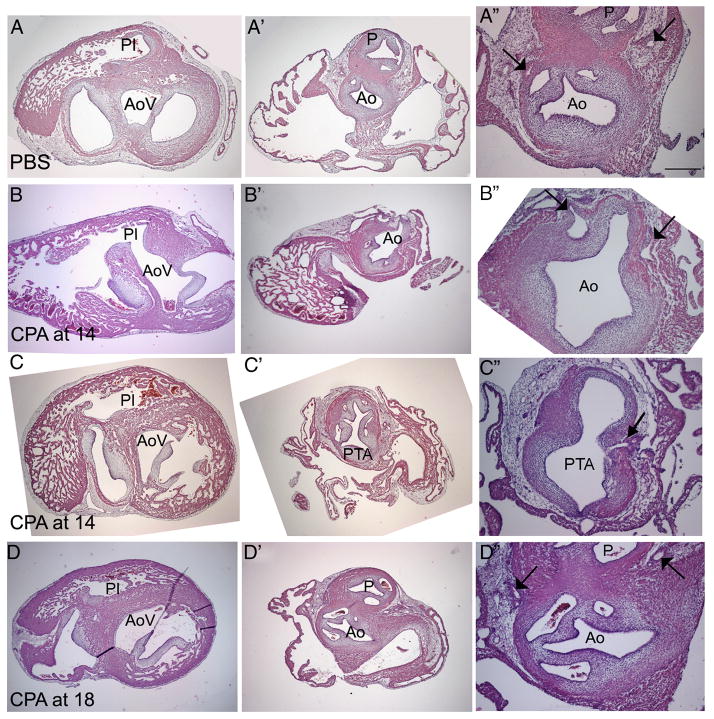
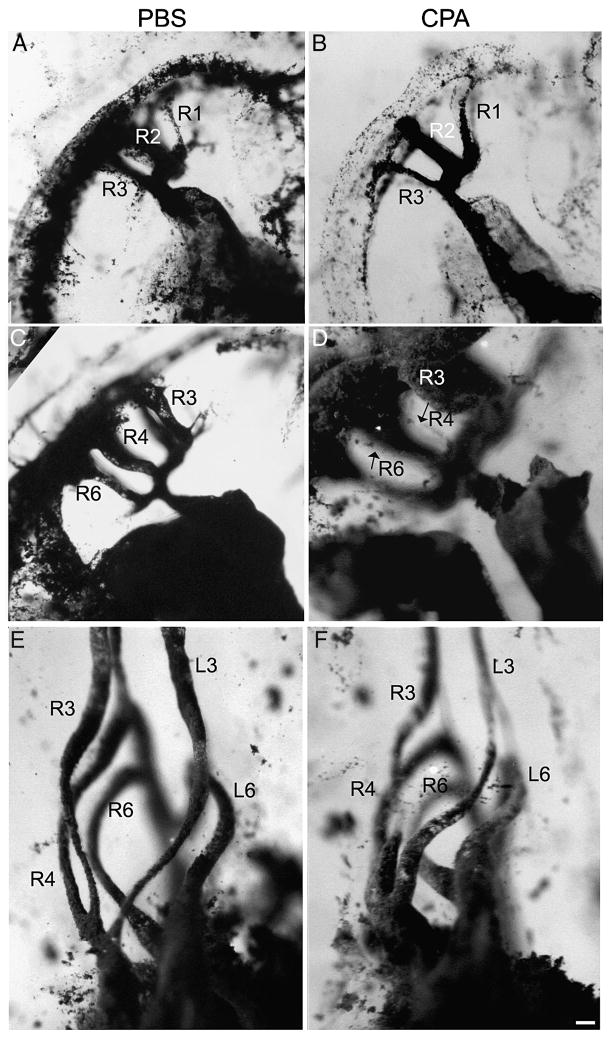
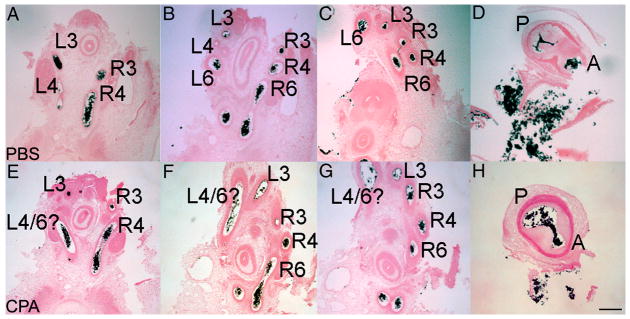
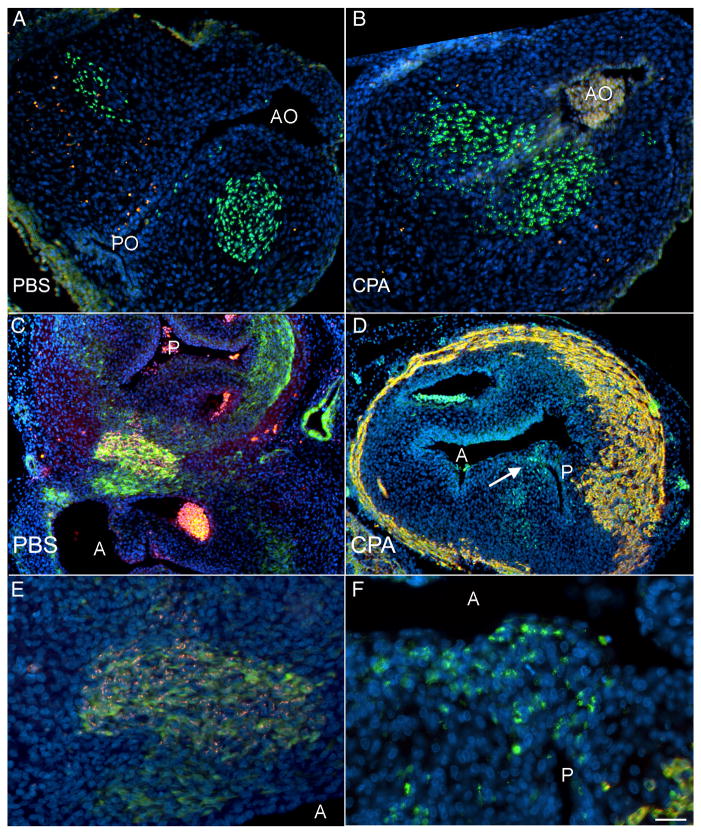
The Sonic hedgehog (Shh)-null mouse was initially described as a phenotypic mimic of Tetralogy of Fallot with pulmonary atresia (Washington Smoak, I., Byrd, N.A., Abu-Issa, R., Goddeeris, M.M., Anderson, R., Morris, J., Yamamura, K., Klingensmith, J., and Meyers, E.N. 2005. Sonic hedgehog is required for cardiac outflow tract and neural crest cell development. Dev. Biol. 283, 357-372.); however, subsequent reports describe only a single outflow tract, leaving the phenotype and its developmental mechanism unclear. We hypothesized that the phenotype that occurs in response to Shh knockdown is pulmonary atresia and is directly related to the abnormal development of the secondary heart field. We found that Shh was expressed by the pharyngeal endoderm adjacent to the secondary heart field and that its receptor Ptc2 was expressed in a gradient in the secondary heart field, with the most robust expression in the caudal secondary heart field, closest to the Shh expression. In vitro culture of secondary heart field with the hedgehog inhibitor cyclopamine significantly reduced proliferation. In ovo, cyclopamine treatment before the secondary heart field adds to the outflow tract reduced proliferation only in the caudal secondary heart field, which coincided with the region of high Ptc2 expression. After outflow tract septation should occur, embryos treated with cyclopamine exhibited pulmonary atresia, pulmonary stenosis, and persistent truncus arteriosus. In hearts with pulmonary atresia, cardiac neural crest-derived cells, which form the outflow tract septum, migrated into the outflow tract and formed a septum. However, this septum divided the outflow tract into two unequal sized vessels and effectively closed off the pulmonary outlet. These experiments show that Shh is necessary for secondary heart field proliferation, which is required for normal pulmonary trunk formation, and that embryos with pulmonary atresia have an outflow tract septum.
Figures


Similar articles
-
BMP signaling modulates hedgehog-induced secondary heart field proliferation.Dev Biol. 2010 Dec 15;348(2):167-76. doi: 10.1016/j.ydbio.2010.09.021. Epub 2010 Oct 14. Dev Biol. 2010. PMID: 20920499 Free PMC article.
-
sonic hedgehog is required in pulmonary endoderm for atrial septation.Development. 2009 May;136(10):1761-70. doi: 10.1242/dev.034157. Epub 2009 Apr 15. Development. 2009. PMID: 19369393 Free PMC article.
-
Independent requirements for Hedgehog signaling by both the anterior heart field and neural crest cells for outflow tract development.Development. 2007 Apr;134(8):1593-604. doi: 10.1242/dev.02824. Epub 2007 Mar 7. Development. 2007. PMID: 17344228
-
Sonic hedgehog is required for cardiac outflow tract and neural crest cell development.Dev Biol. 2005 Jul 15;283(2):357-72. doi: 10.1016/j.ydbio.2005.04.029. Dev Biol. 2005. PMID: 15936751
-
The role of secondary heart field in cardiac development.Dev Biol. 2009 Dec 15;336(2):137-44. doi: 10.1016/j.ydbio.2009.10.009. Epub 2009 Oct 14. Dev Biol. 2009. PMID: 19835857 Free PMC article. Review.
Cited by
-
The Classification of VACTERL Association into 3 Groups According to the Limb Defect.Plast Reconstr Surg Glob Open. 2021 Feb 2;9(2):e3360. doi: 10.1097/GOX.0000000000003360. eCollection 2021 Feb. Plast Reconstr Surg Glob Open. 2021. PMID: 33680640 Free PMC article.
-
A Membrane-Tethered Ubiquitination Pathway Regulates Hedgehog Signaling and Heart Development.Dev Cell. 2020 Nov 23;55(4):432-449.e12. doi: 10.1016/j.devcel.2020.08.012. Epub 2020 Sep 22. Dev Cell. 2020. PMID: 32966817 Free PMC article.
-
A 3 base pair deletion in TBX1 leads to reduced protein expression and transcriptional activity.Sci Rep. 2017 Mar 8;7:44165. doi: 10.1038/srep44165. Sci Rep. 2017. PMID: 28272434 Free PMC article. Clinical Trial.
-
Cilia and coordination of signaling networks during heart development.Organogenesis. 2014 Jan 1;10(1):108-25. doi: 10.4161/org.27483. Epub 2013 Dec 17. Organogenesis. 2014. PMID: 24345806 Free PMC article. Review.
-
Follow Me! A Tale of Avian Heart Development with Comparisons to Mammal Heart Development.J Cardiovasc Dev Dis. 2020 Mar 7;7(1):8. doi: 10.3390/jcdd7010008. J Cardiovasc Dev Dis. 2020. PMID: 32156044 Free PMC article. Review.
References
-
- Abu-Issa R, Smyth G, Smoak I, Yamamura K, Meyers EN. Fgf8 is required for pharyngeal arch and cardiovascular development in the mouse. Development. 2002;129:4613–25. - PubMed
-
- Allan GJ, Zannoni A, McKinnel I, Otto WR, Holzenberger M, Flint DJ, Patel K. Major components of the insulin-like growth factor axis are expressed early in chicken embryogenesis, with IGF binding protein (IGFBP)-5 expression subject to regulation by Sonic Hedgehog. Anat Embryol (Berl) 2003;207:73–84. - PubMed
-
- Alvarez-Rodriguez R, Barzi M, Berenguer J, Pons S. Bone morphogenetic protein 2 opposes Shh-mediated proliferation in cerebellar granule cells through a TIEG-1-based regulation of Nmyc. J Biol Chem. 2007;282:37170–37180. - PubMed
-
- Arnold JS, Werling U, Braunstein EM, Liao J, Nowotschin S, Edelmann W, Hebert JM, Morrow BE. Inactivation of Tbx1 in the pharyngeal endoderm results in 22q11DS malformations. Development. 2006;133:977–987. - PubMed
-
- Bajolle F, Zaffran S, Meilhac SM, Dandonneau M, Chang T, Kelly RG, Buckingham ME. Myocardium at the base of the aorta and the pulmonary trunk is prefigured in the outflow tract of the heart and in subdomains of the second heart field. Dev Biol. 2008;313:25–34. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
