

The intracellular sensor NLRP3 mediates key innate and healing responses to influenza A virus via the regulation of caspase-1
- PMID: 19362023
- PMCID: PMC2765464
- DOI: 10.1016/j.immuni.2009.02.006
The intracellular sensor NLRP3 mediates key innate and healing responses to influenza A virus via the regulation of caspase-1
Abstract
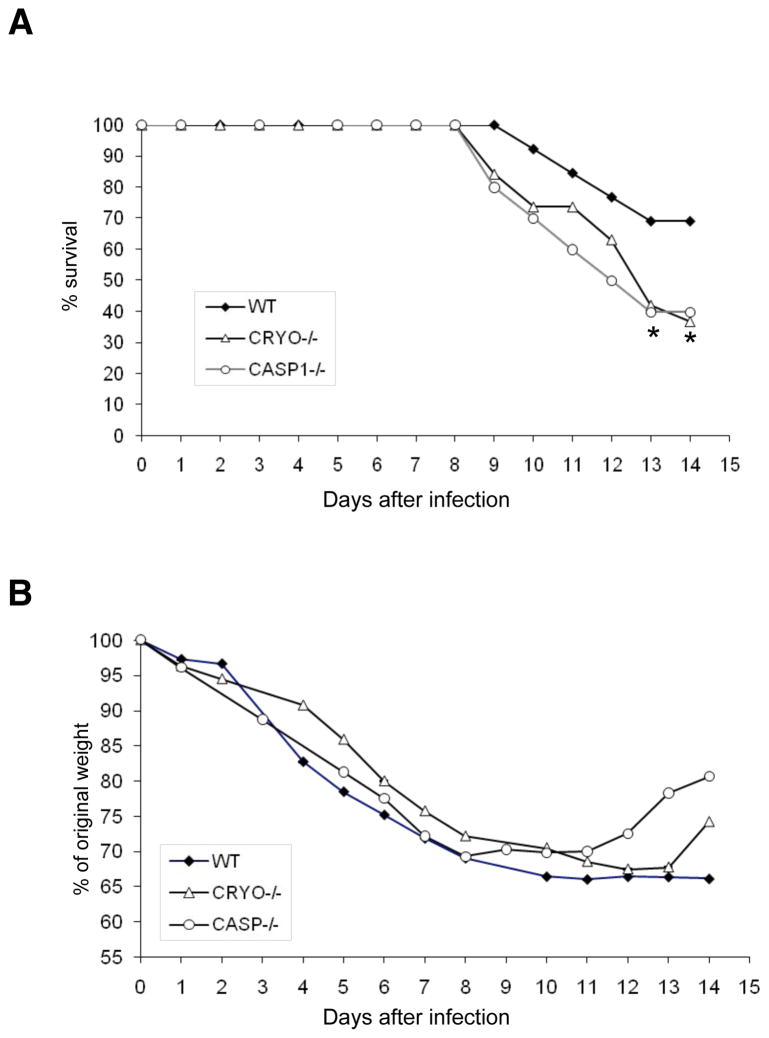
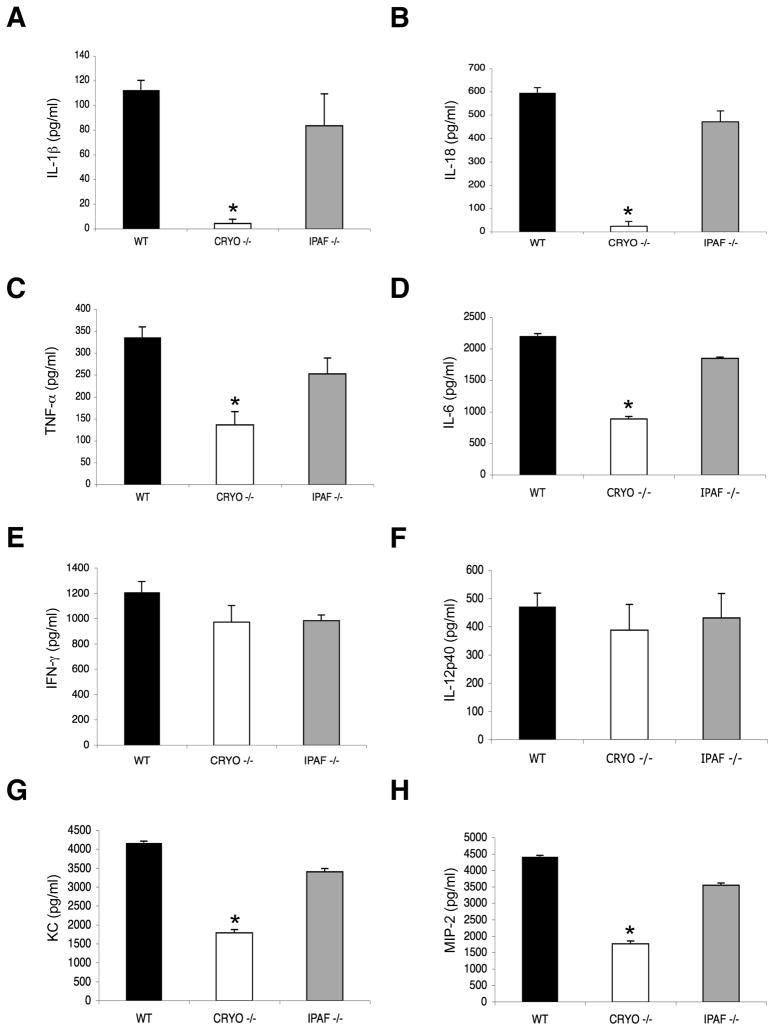
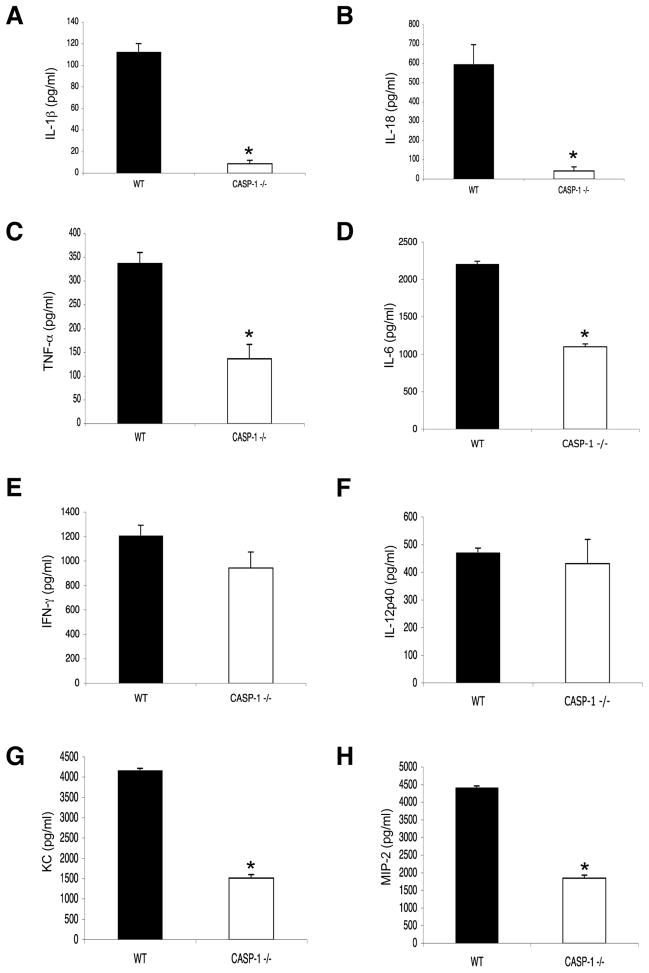
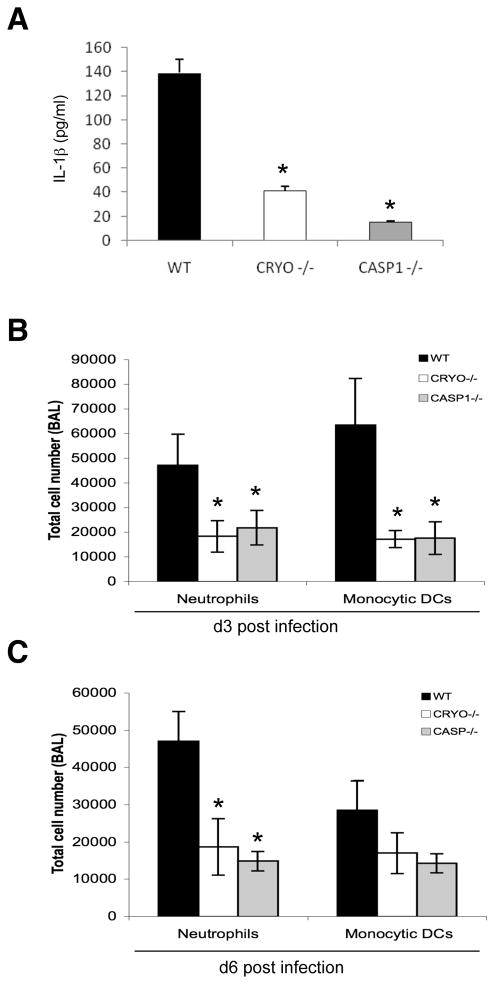
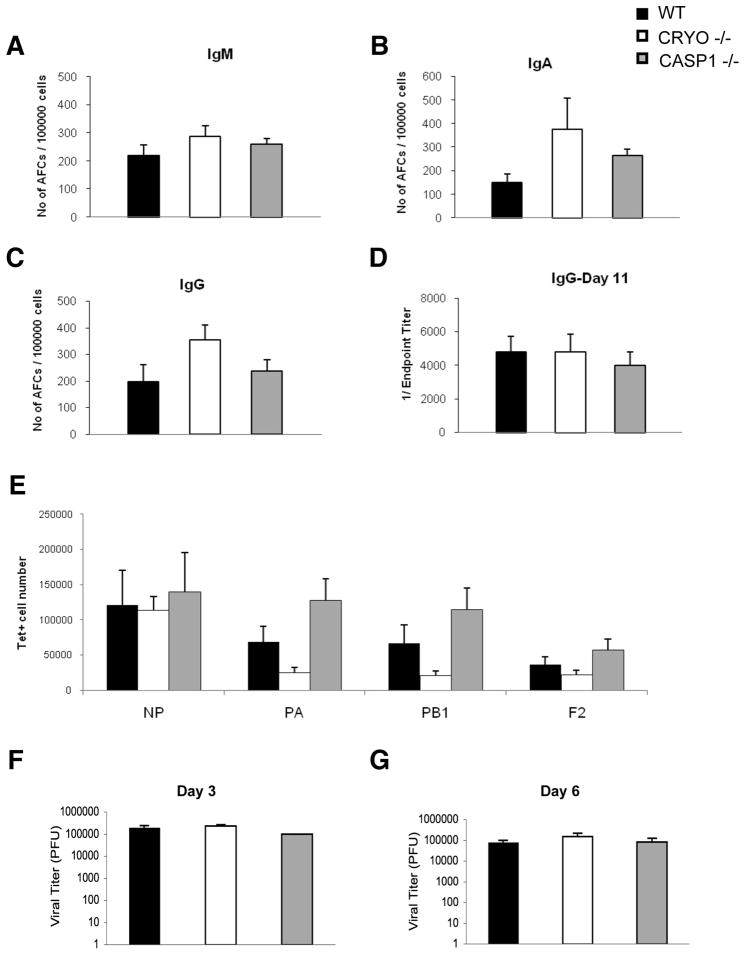
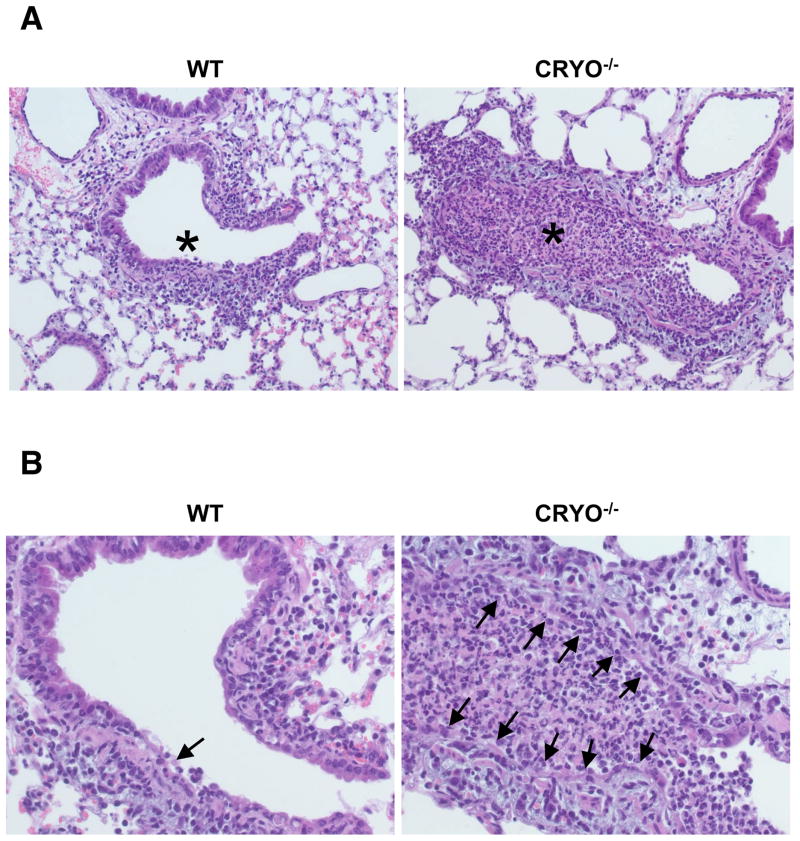
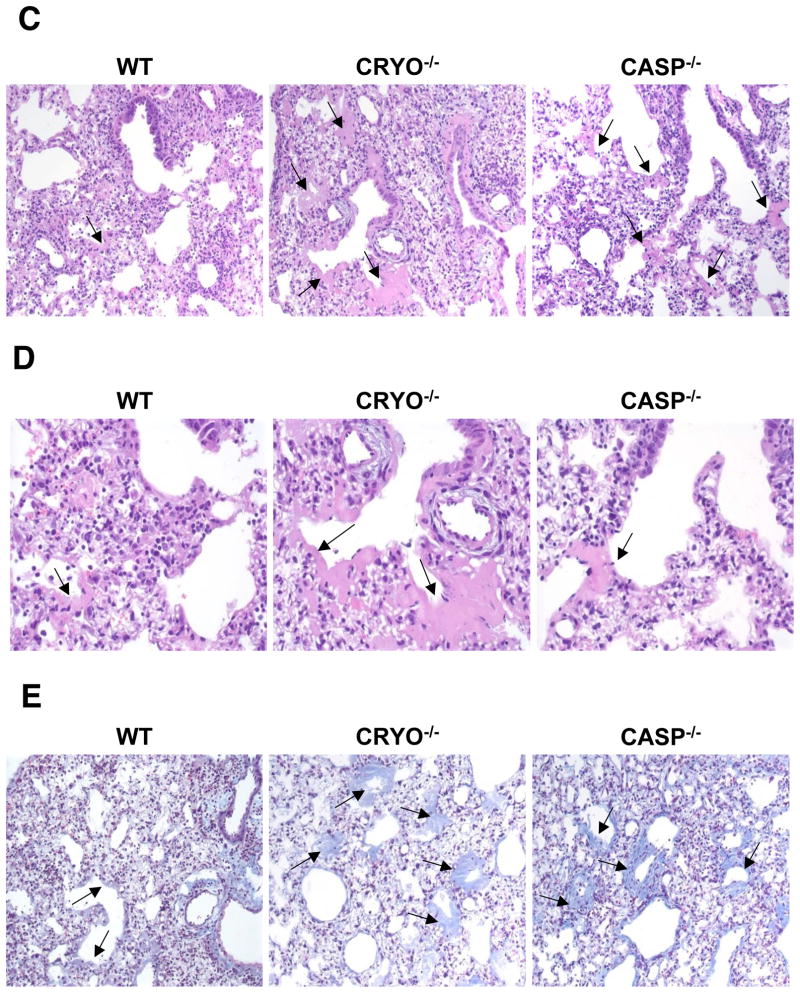
Virus-induced interlukin-1beta (IL-1beta) and IL-18 production in macrophages are mediated via caspase-1 pathway. Multiple microbial components, including viral RNA, are thought to trigger assembly of the cryopyrin inflammasome resulting in caspase-1 activation. Here, we demonstrated that Nlrp3(-/-) and Casp1(-/-) mice were more susceptible than wild-type mice after infection with a pathogenic influenza A virus. This enhanced morbidity correlated with decreased neutrophil and monocyte recruitment and reduced cytokine and chemokine production. Despite the effect on innate immunity, cryopyrin-deficiency was not associated with any obvious defect in virus control or on the later emergence of the adaptive response. Early epithelial necrosis was, however, more severe in the infected mutants, with extensive collagen deposition leading to later respiratory compromise. These findings reveal a function of the cryopyrin inflammasome in healing responses. Thus, cryopyrin and caspase-1 are central to both innate immunity and to moderating lung pathology in influenza pneumonia.
Figures
References
-
- Allan W, Tabi Z, Cleary A, Doherty PC. Cellular events in the lymph node and lung of mice with influenza. Consequences of depleting CD4+ T cells. J Immunol. 1990;144:3980–3986. - PubMed
-
- Amer A, Franchi L, Kanneganti TD, Body-Malapel M, Ozoren N, Brady G, Meshinchi S, Jagirdar R, Gewirtz A, Akira S, et al. Regulation of Legionella phagosome maturation and infection through flagellin and host IPAF. J Biol Chem 2006 - PubMed
-
- Burns K, Martinon F, Tschopp J. New insights into the mechanism of IL-1beta maturation. Curr Opin Immunol. 2003;15:26–30. - PubMed
-
- Denton AE, Doherty PC, Turner SJ, La Gruta NL. IL-18, but not IL-12, is required for optimal cytokine production by influenza virus-specific CD8+ T cells. Eur J Immunol. 2007;37:368–375. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
