

Toward a structural understanding of IRES RNA function
- PMID: 19362464
- PMCID: PMC2757110
- DOI: 10.1016/j.sbi.2009.03.005
Toward a structural understanding of IRES RNA function
Abstract
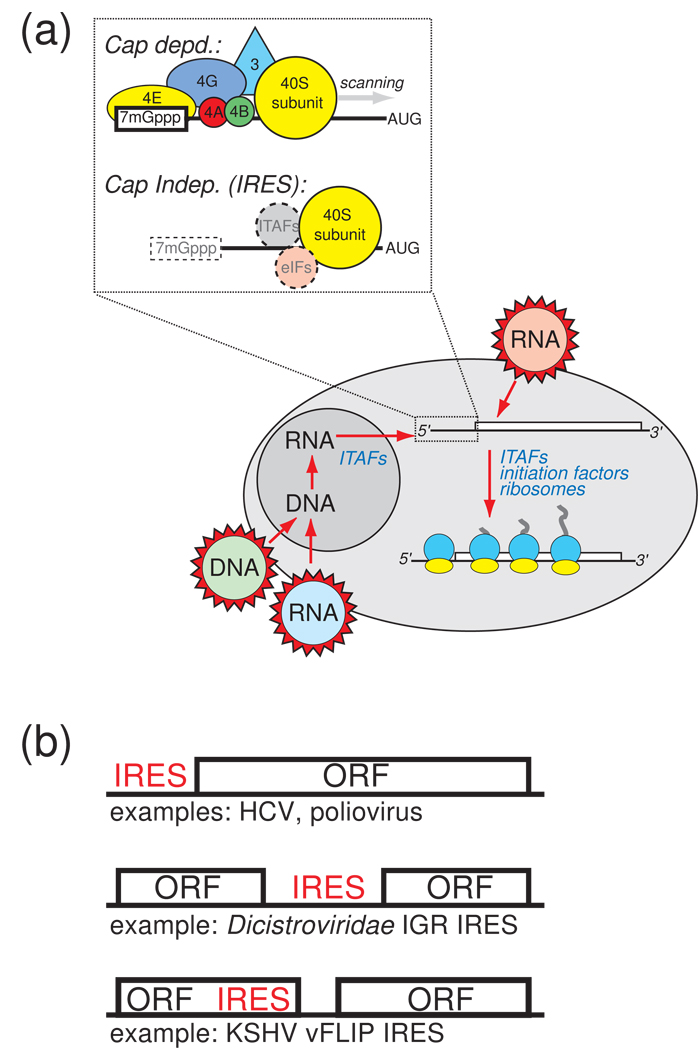
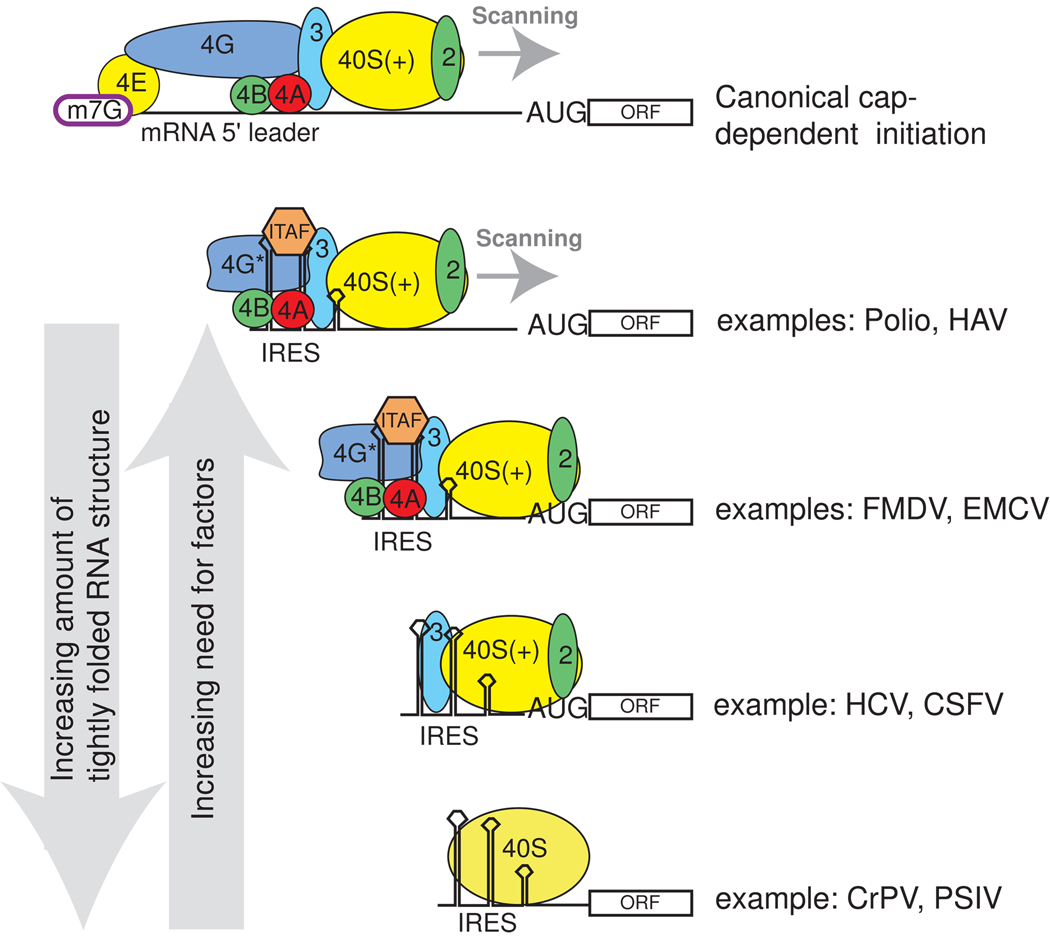
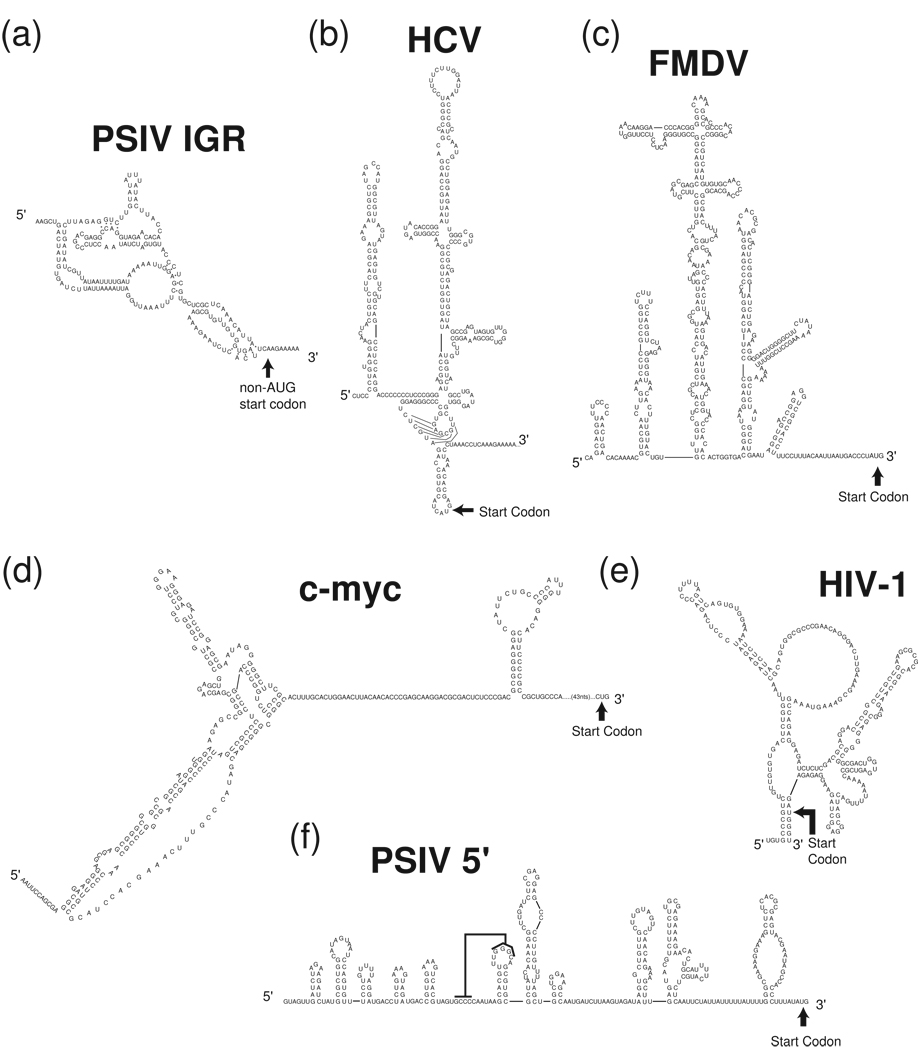
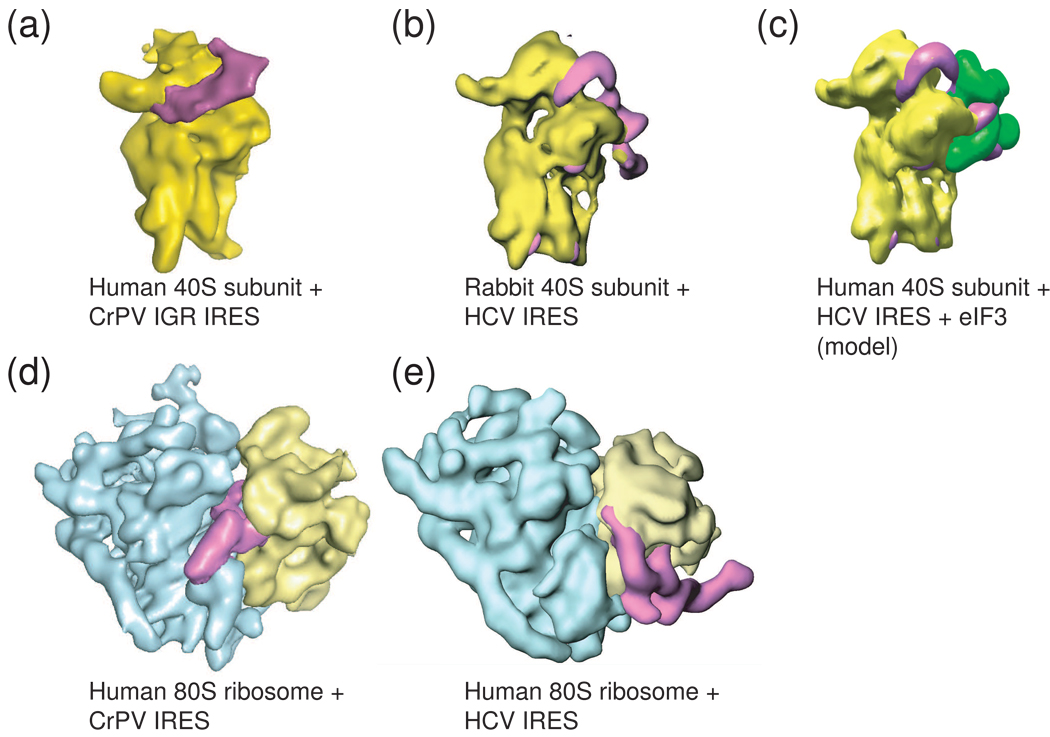
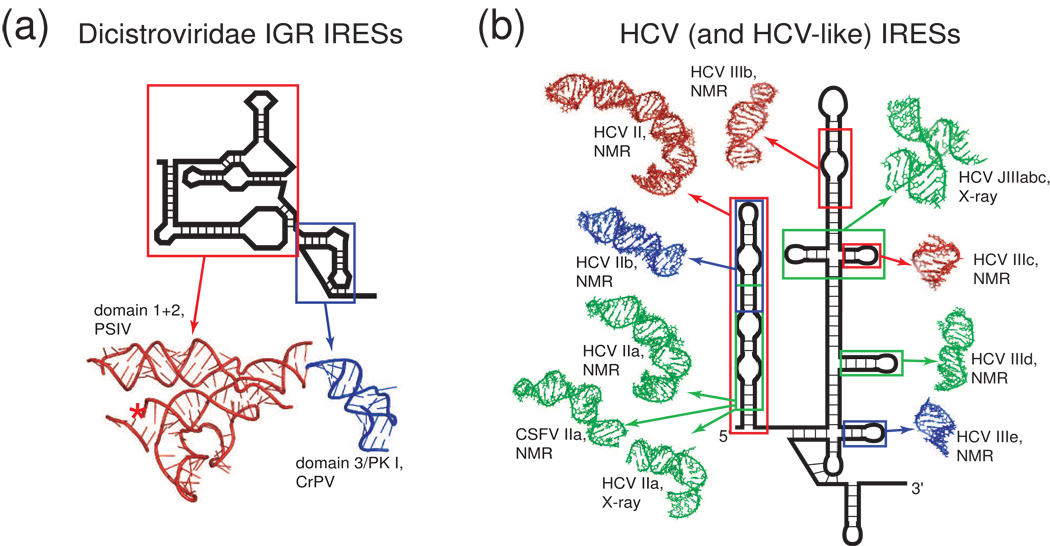
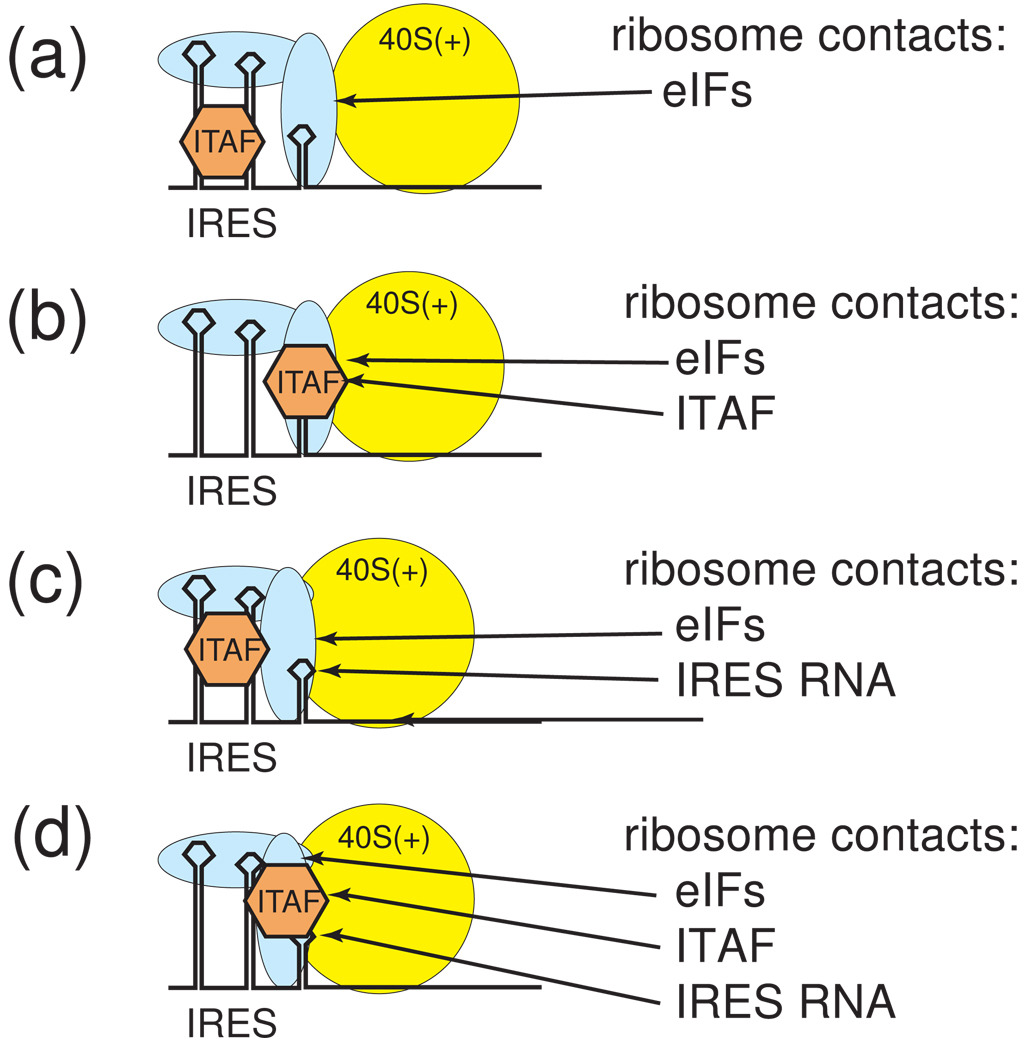
Protein synthesis of an RNA template can start by two different known mechanisms: cap-dependent translation initiation and cap-independent translation initiation. The latter is driven by RNA sequences called internal ribosome entry sites (IRESs) that are found in both viral RNAs and cellular mRNAs. The diverse mechanisms used by IRESs are reflected in their structural diversity, and this structural diversity challenges us to develop a cohesive model linking IRES function to structure. With more direct structural information available for the viral IRESs, data suggest an inverse correlation between the degree to which an IRES RNA can form a stable structure on its own and the number of factors that it requires to function. Lessons learned from the viral IRESs may help understand the cellular IRESs, although more structural data are needed before any strong links can be made.
Figures
References
-
- Pestova T, Lorsch JR, Hellen CU. The Mechanism of Translation Initiation in Eukaryotes. In: Mathews MB, Sonenberg N, editors. Translational Control in Biology and Medicine. Hershey JWB: Cold Sring Harbor Laboratory Press; 2007. pp. 87–128.
-
- Doudna JA, Sarnow P. Translation initiation by viral internal ribosome entry sites. In: Mathews MB, Sonenberg N, editors. Translational Control in Biology and Medicine. Hershey JWB: Cold Spring Harbor Laboratory Press; 2007. pp. 129–153.
-
- Elroy-Stein O, Merrick WC. Translation initiation via cellular internal ribosome entry sites. In: Mathews MB, Sonenberg N, Hershey J, editors. Translational Control in Biology and Medicine. Hershey J: Cold Spring Harbor Laboratroy Press; 2007. pp. 155–172.
-
- Jackson RJ. Alternative mechanisms of initiating translation of mammalian mRNAs. Biochem Soc Trans. 2005;33:1231–1241. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
