

doi: 10.1016/S0076-6879(09)04617-5.
Bacterial fatty acid synthesis and its relationships with polyketide synthetic pathways
Affiliations
- PMID: 19362649
- PMCID: PMC4095770
- DOI: 10.1016/S0076-6879(09)04617-5
Item in Clipboard
Bacterial fatty acid synthesis and its relationships with polyketide synthetic pathways
Methods Enzymol.
2009.
Abstract
This review presents the most thoroughly studied bacterial fatty acid synthetic pathway, that of Escherichia coli and then discusses the exceptions to the E. coli pathway present in other bacteria. The known interrelationships between the fatty acid and polyketide synthetic pathways are also assessed, mainly in the Streptomyces group of bacteria. Finally, we present a compendium of methods for analysis of bacterial fatty acid synthetic pathways.
Figures

Typical bacterial fatty acids. Structure A is hexadecanoic (palmitic) acid, a very abundant saturated fatty acid whereas structure B is cis-9-hexadecenoic (palmitoleic) acid which together with the product formed by one additional spin of the elongation cycle, cis-11-octadecenoic (cis-vaccenic) acid, are the dominant unsaturated species found in bacteria. Structure C is the cyclopropane fatty acid cis-9,10-methylene hexadecanoic acid formed by methylene incorporation from S-adenosyl-L-methionine into phospholipid-bound palmitoleic acid. This together with its C19 homologue are distributed very widely in bacteria. Structures D and E are C15 branched chain fatty acids where D is the anteiso species and E is the iso species. Structure F is 3-hydroxytetadecanoic acid, a major component of the lipid A of most gram negative bacteria.

ACP prosthetic group metabolism in E. coli. Apo-ACP, the product of the acpP gene, is non-functional in fatty acid synthesis. It is post-translationally activated by the attachment of a 4’phosphopantetheine arm derived from coenzyme A, a reaction catalyzed by the holo-ACP synthase AcpS. Turnover of ACP involves removal of the prosthetic group by ACP hydrolyase, AcpH which can then be used in the synthesis of CoA.

The initiation steps in the type II fatty acid synthesis pathway of E. coli. Malonyl-CoA is converted to malonyl-ACP by malonyl transacylase (FabD). Fatty acid synthesis is initiated by FabH, which condenses malonyl-ACP with acetyl-CoA.
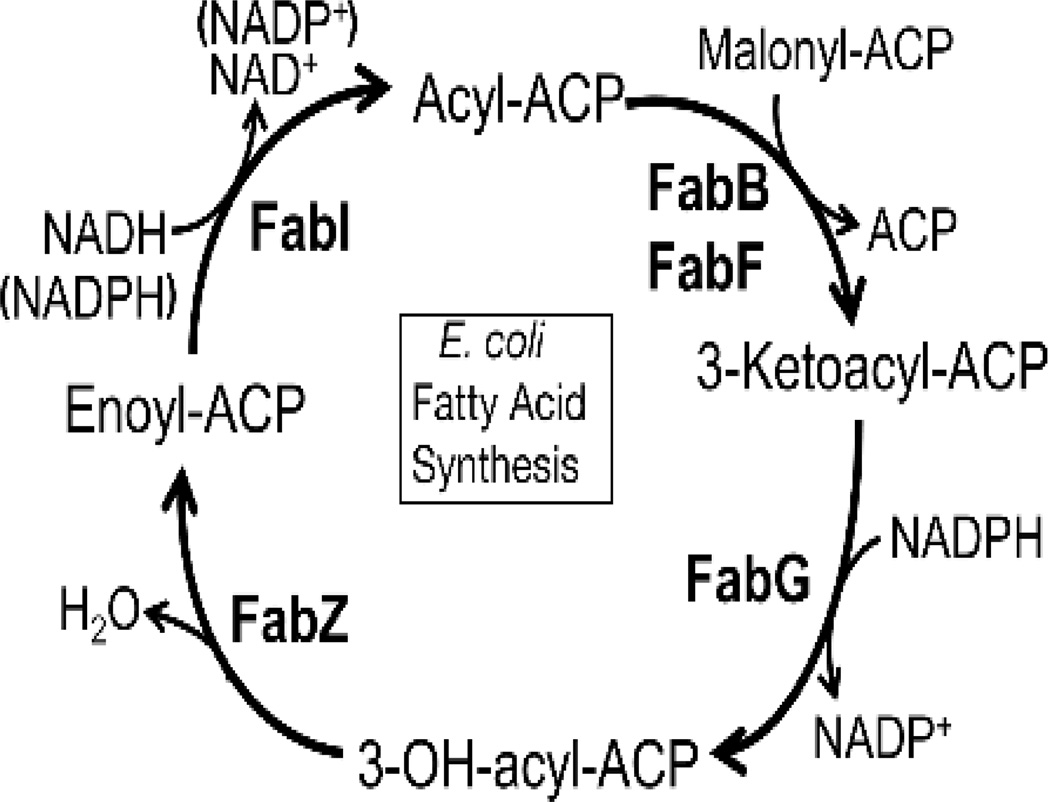
The elongation cycle. There are four steps in fatty acid elongation. Each new cycle of 2-carbon elongation is initiated by the condensation of acyl-ACP and malonyl-ACP by one of the elongation condensing enzymes, FabB or FabF. The next step is the reduction of the 3-ketoacyl-ACP by the NADPH-dependent FabG. This 3-hydroxyacyl-ACP is dehydrated to trans-2-acyl-ACP by FabZ. The final step is the NADH-dependent reduction of enoyl-ACP to acyl-ACP by FabI.

Production of unsaturated fatty acids. Unsaturated fatty acids arise from a branch in the biosynthetic pathway at the 3-hydroxydecanoyl-ACP intermediate. FabA is a unique 3-hydroxyacyl-ACP dehydratase that is capable for forming trans-2-decenoyl-ACP and isomerizing this intermediate to cis-3-decenoyl-ACP. FabA is very selective for the 10-carbon substrates in vivo. FabB is absolutely required for the subsequent elongation of cis-3-decenoyl-ACP to 16:1-ACP. Interestingly, 16:1-ACP is a poor substrate for FabB, and its elongation to 18:1-ACP is controlled by the activity of FabF. FabF is a naturally temperature-sensitive enzyme, and this property account for the greater proportion of 18:1 in bacteria grown at low temperatures compared to those grown at higher temperatures. The trans-3-decenoyl-ACP is used by FabI followed by elongation by either FabB or FabF to form palmitic acid, the major saturated fatty acid in E. coli. The parentheses show isozymes from other bacteria that catalyze the reaction shown.
References
-
- Alberts AA, Majerus PW, Vagelos PR. Malonyl-CoA acyl carrier protein transacylase. Meth. Enzymol. 1974;14:53–56.
-
- Alberts AW, Bell RM, Vagelos PR. Acyl carrier protein. XV. Studies of β-ketoacyl-acyl carrier protein synthetase. J. Biol. Chem. 1972;247:3190–3198. - PubMed
-
- Arthur CJ, Szafranska A, Evans SE, Findlow SC, Burston SG, Owen P, Clark-Lewis I, Simpson TJ, Crosby J, Crump MP. Self-malonylation is an intrinsic property of a chemically synthesized type II polyketide synthase acyl carrier protein. Biochemistry. 2005;44:15414–15421. - PubMed
-
- Arthur CJ, Szafranska AE, Long J, Mills J, Cox RJ, Findlow SC, Simpson TJ, Crump MP, Crosby J. The malonyl transferase activity of type II polyketide synthase acyl carrier proteins. Chem. Biol. 2006;13:587–596. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
