

Phytochrome B enhances photosynthesis at the expense of water-use efficiency in Arabidopsis
- PMID: 19363093
- PMCID: PMC2689964
- DOI: 10.1104/pp.109.135509
Phytochrome B enhances photosynthesis at the expense of water-use efficiency in Arabidopsis
Abstract
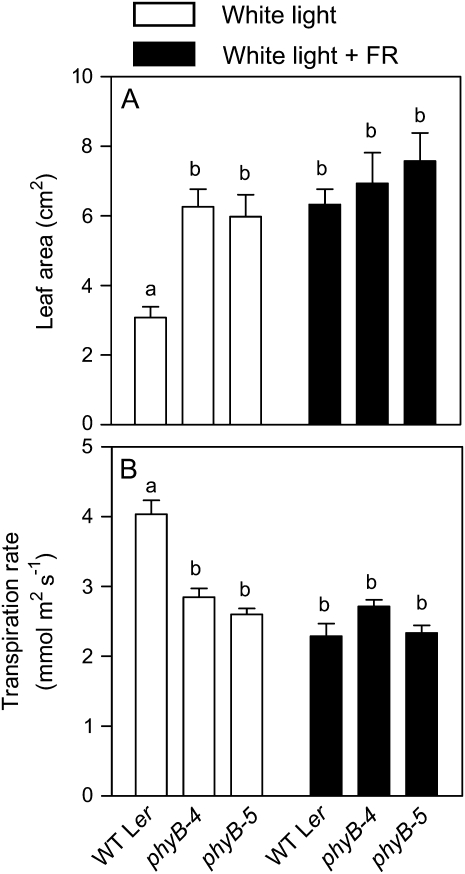
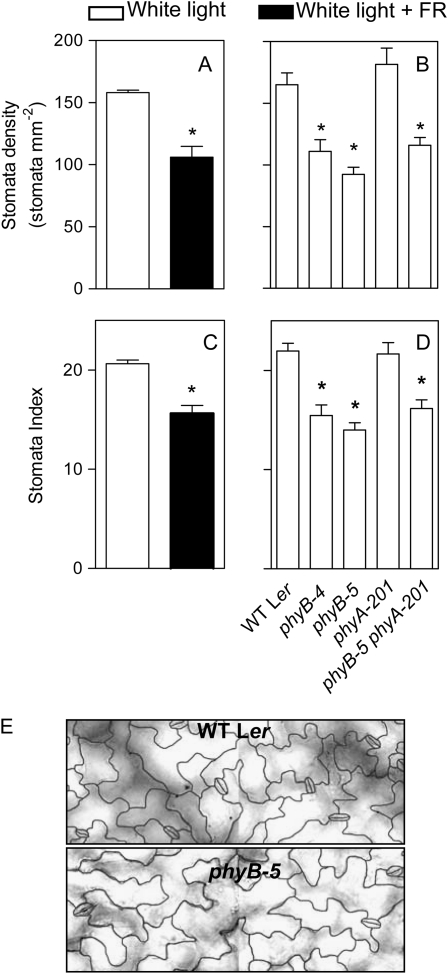
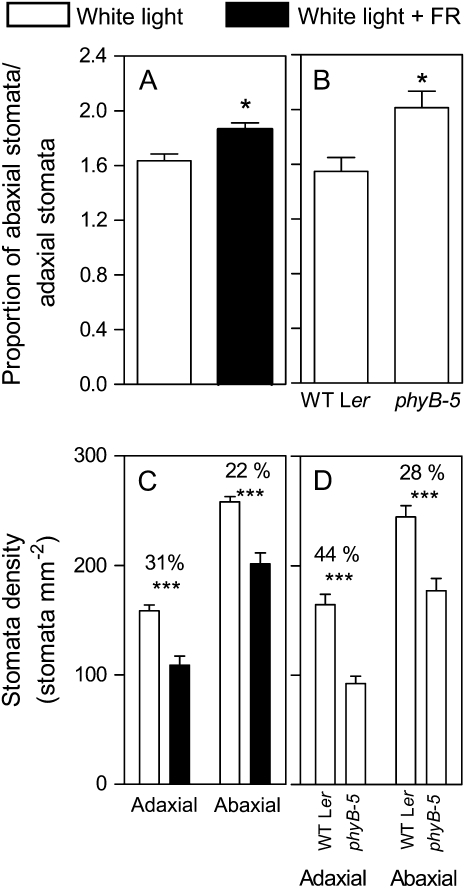
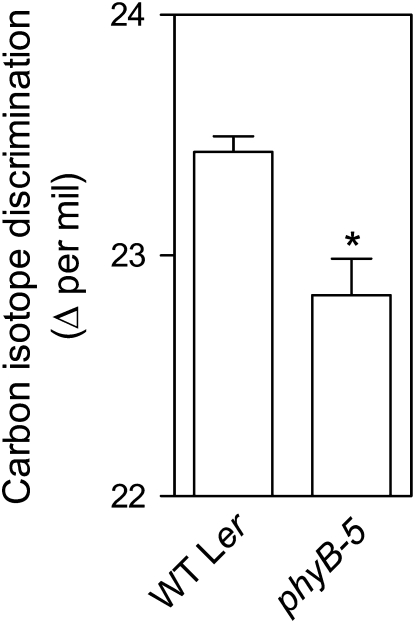
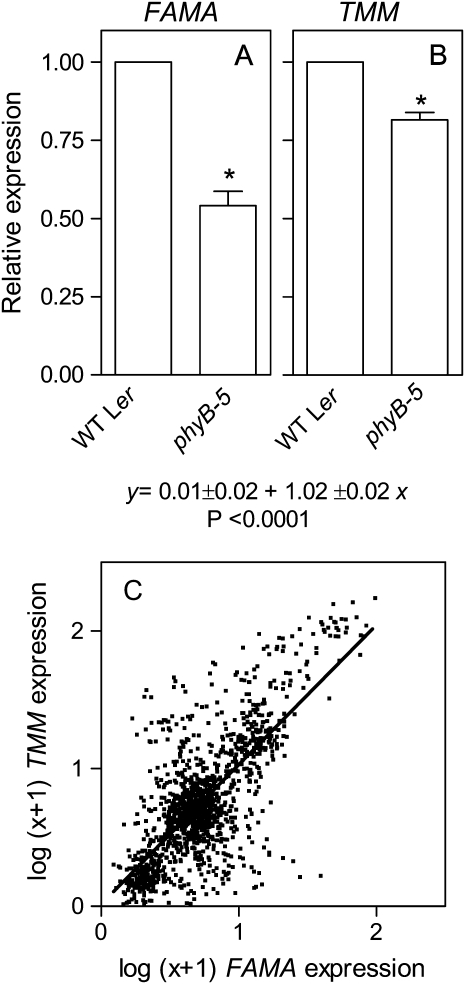
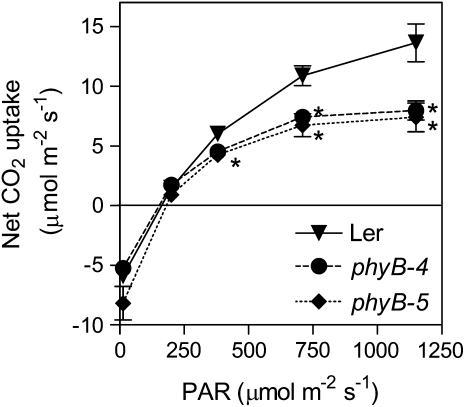
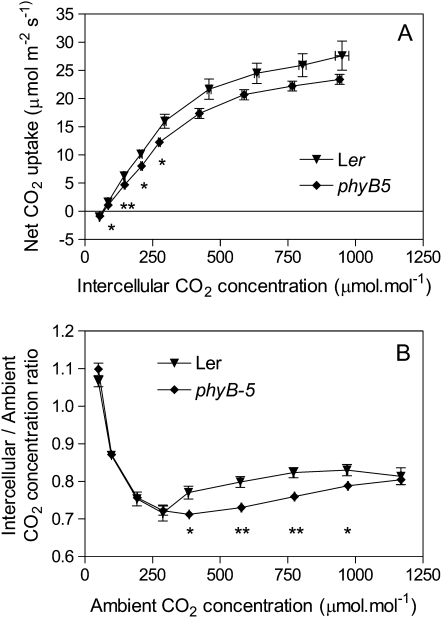
In open places, plants are exposed to higher fluence rates of photosynthetically active radiation and to higher red to far-red ratios than under the shade of neighbor plants. High fluence rates are known to increase stomata density. Here we show that high, compared to low, red to far-red ratios also increase stomata density in Arabidopsis (Arabidopsis thaliana). High red to far-red ratios increase the proportion of phytochrome B (phyB) in its active form and the phyB mutant exhibited a constitutively low stomata density. phyB increased the stomata index (the ratio between stomata and epidermal cells number) and the level of anphistomy (by increasing stomata density more intensively in the adaxial than in the abaxial face). phyB promoted the expression of FAMA and TOO MANY MOUTHS genes involved in the regulation of stomata development in young leaves. Increased stomata density resulted in increased transpiration per unit leaf area. However, phyB promoted photosynthesis rates only at high fluence rates of photosynthetically active radiation. In accordance to these observations, phyB reduced long-term water-use efficiency estimated by the analysis of isotopic discrimination against (13)CO(2). We propose a model where active phyB promotes stomata differentiation in open places, allowing plants to take advantage of the higher irradiances at the expense of a reduction of water-use efficiency, which is compensated by a reduced leaf area.
Figures
References
-
- Aphalo PJ, Gibson D, Di Benedetto AH (1991) Responses of growth, photosynthesis, and leaf conductance to white light irradiance and end-of-day red and far-red pulses in Fuchsia magellanica Lam. New Phytol 117 461–471 - PubMed
-
- Assmann SM (1992) Effects of light quantity and quality during development on the morphology and stomatal physiology of Commelina communis. Oecologia 92 188–195 - PubMed
-
- Ballaré CL, Sánchez RA, Scopel AL, Casal JJ, Ghersa CM (1987) Early detection of neighbour plants by phytochrome perception of spectral changes in reflected sunlight. Plant Cell Environ 10 551–557
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
