

Intersection of two signalling pathways: extracellular nucleotides regulate pollen germination and pollen tube growth via nitric oxide
- PMID: 19363208
- PMCID: PMC2682505
- DOI: 10.1093/jxb/erp091
Intersection of two signalling pathways: extracellular nucleotides regulate pollen germination and pollen tube growth via nitric oxide
Abstract
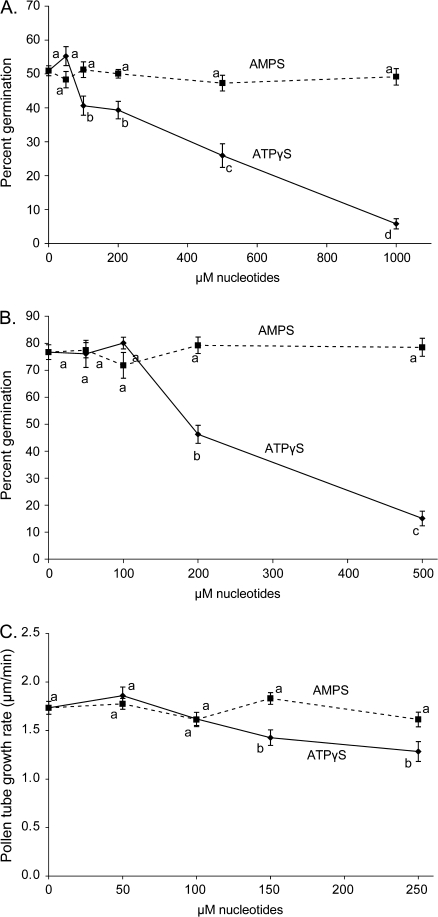
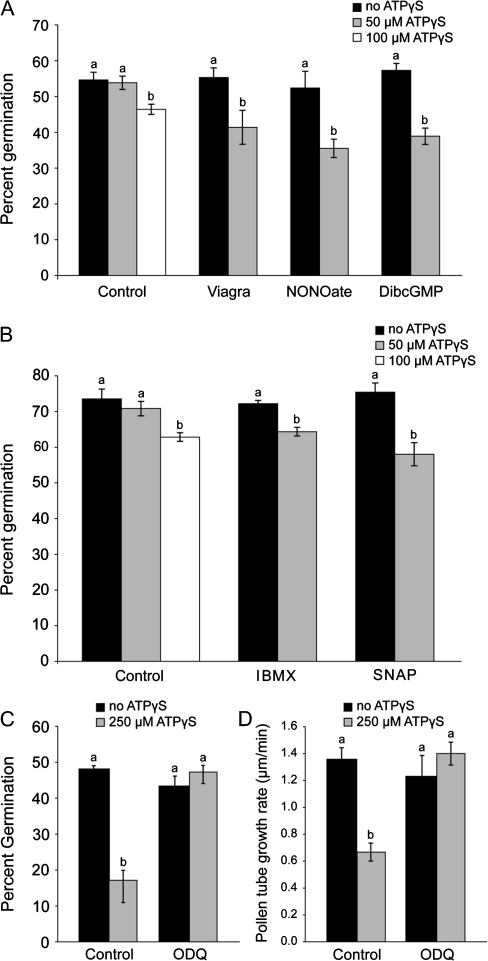
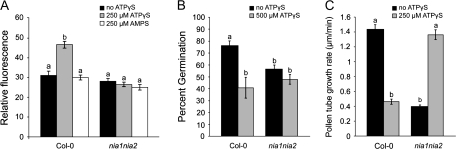
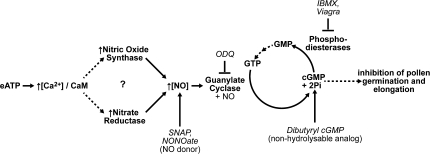
Plant and animal cells release or secrete ATP by various mechanisms, and this activity allows extracellular ATP to serve as a signalling molecule. Recent reports suggest that extracellular ATP induces plant responses ranging from increased cytosolic calcium to changes in auxin transport, xenobiotic resistance, pollen germination, and growth. Although calcium has been identified as a secondary messenger for the extracellular ATP signal, other parts of this signal transduction chain remain unknown. Increasing the extracellular concentration of ATPgammaS, a poorly-hydrolysable ATP analogue, inhibited both pollen germination and pollen tube elongation, while the addition of AMPS had no effect. Because pollen tube elongation is also sensitive to nitric oxide, this raised the possibility that a connection exists between the two pathways. Four approaches were used to test whether the germination and growth effects of extracellular ATPgammaS were transduced via nitric oxide. The results showed that increases in extracellular ATPgammaS induced increases in cellular nitric oxide, chemical agonists of the nitric oxide signalling pathway lowered the threshold of extracellular ATPgammaS that inhibits pollen germination, an antagonist of guanylate cyclase, which can inhibit some nitric oxide signalling pathways, blocked the ATPgammaS-induced inhibition of both pollen germination and pollen tube elongation, and the effects of applied ATPgammaS were blocked in nia1nia2 mutants, which have diminished NO production. The concurrence of these four data sets support the conclusion that the suppression of pollen germination and pollen tube elongation by extracellular nucleotides is mediated in part via the nitric oxide signalling pathway.
Figures
References
-
- Besson-Bard A, Pugin A, Wendehenne D. New insights into nitric oxide signalling in plants. Annual Review of Plant Biology. 2008;59:21–39. - PubMed
-
- Beligni M, Lamattina L. Nitric oxide stimulates seed germination and de-etiolation, and inhibits hypocotyl elongation, three light-inducible responses in plants. Planta. 2000;210:215–221. - PubMed
-
- Boavida LC, McCormick S. Temperature as a determinant factor for increased and reproducible in vitro pollen germination in Arabidopsis thaliana. The Plant Journal. 2007;52:570–582. - PubMed
-
- Borisjuk L, Rolletschek H, Walenta S, Panitz R, Wobus U, Weber H. Energy status and its control on embryogenesis of legumes, ATP distribution within Vicia faba embryos is developmentally regulated and correlated with photosynthetic capacity. The Plant Journal. 2003;36:318–330. - PubMed
-
- Bright J, Desikan R, Hancock J, Weir I, Neill S. ABA-induced NO generation and stomatal closure in Arabidopsis are dependent on HP2O2 synthesis. The Plant Journal. 2006;45:113–122. - PubMed
