

The effects of extracellular calcium on motility, pseudopod and uropod formation, chemotaxis, and the cortical localization of myosin II in Dictyostelium discoideum
- PMID: 19363786
- PMCID: PMC2747089
- DOI: 10.1002/cm.20367
The effects of extracellular calcium on motility, pseudopod and uropod formation, chemotaxis, and the cortical localization of myosin II in Dictyostelium discoideum
Abstract
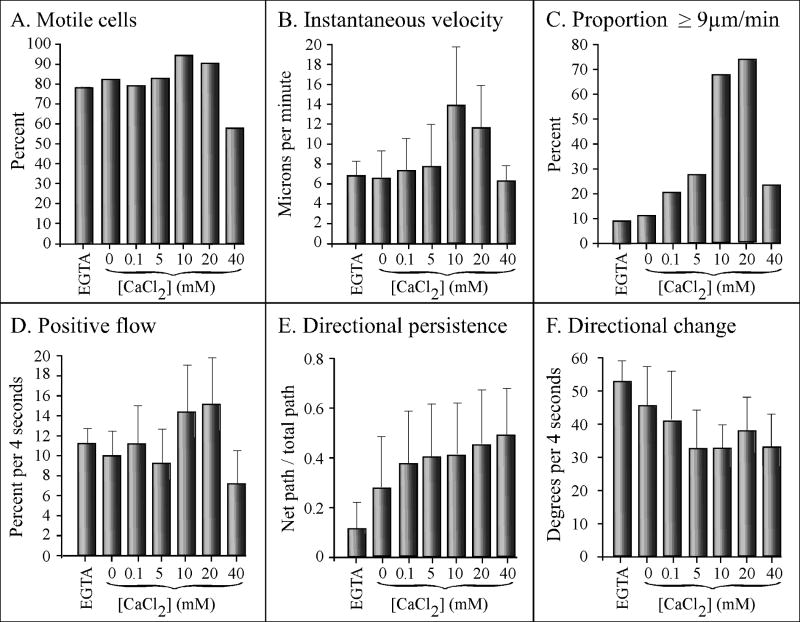
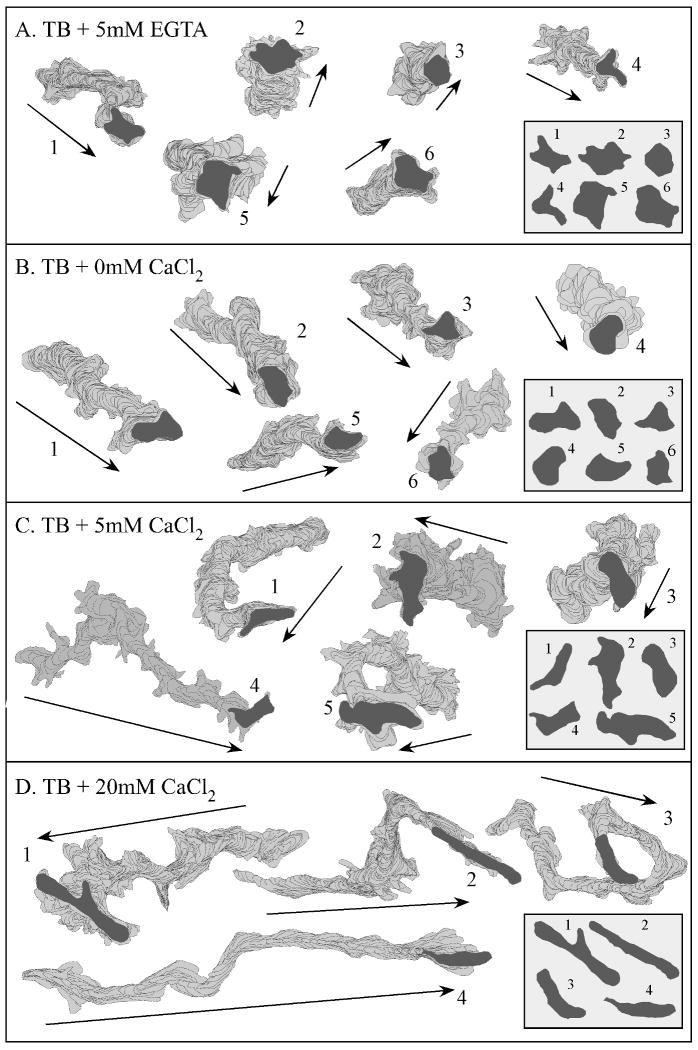
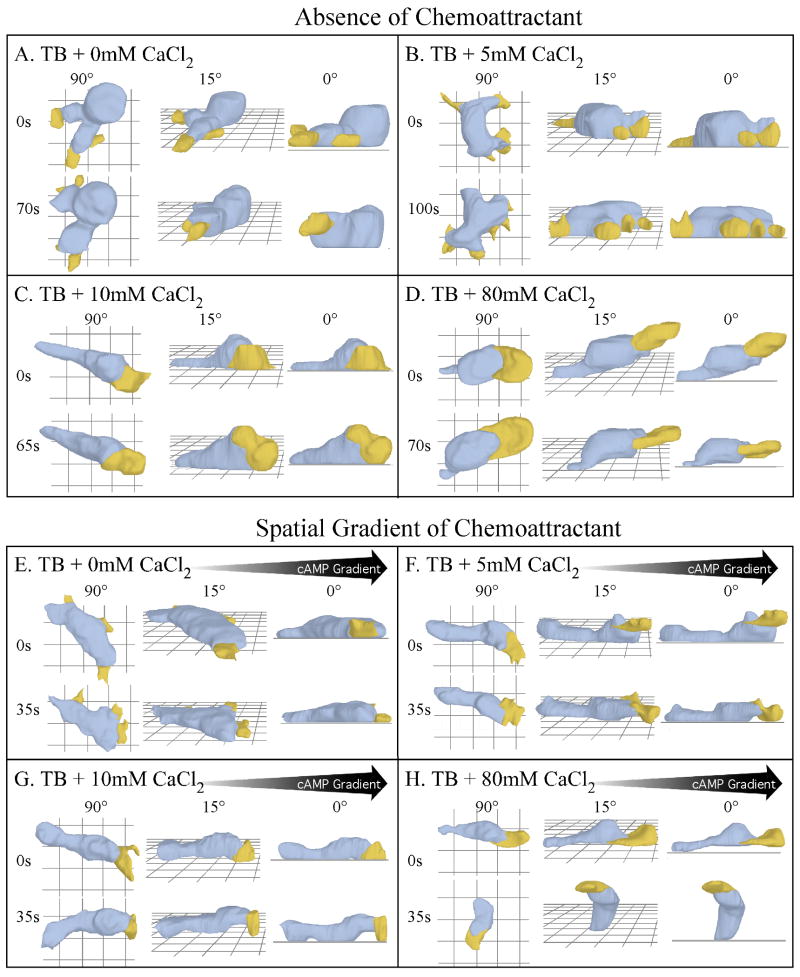
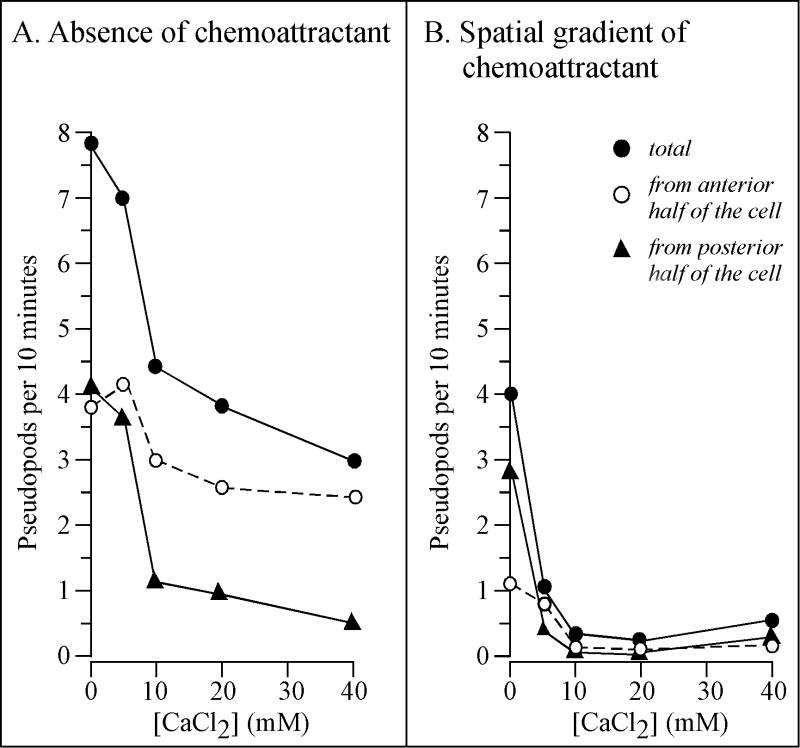
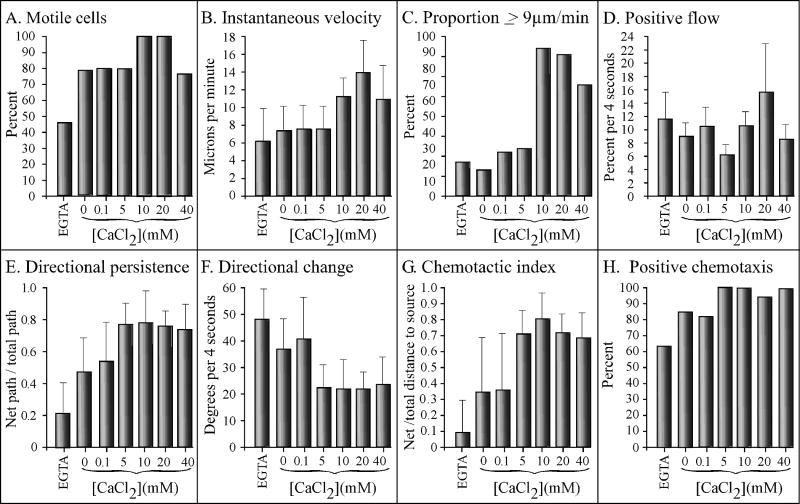
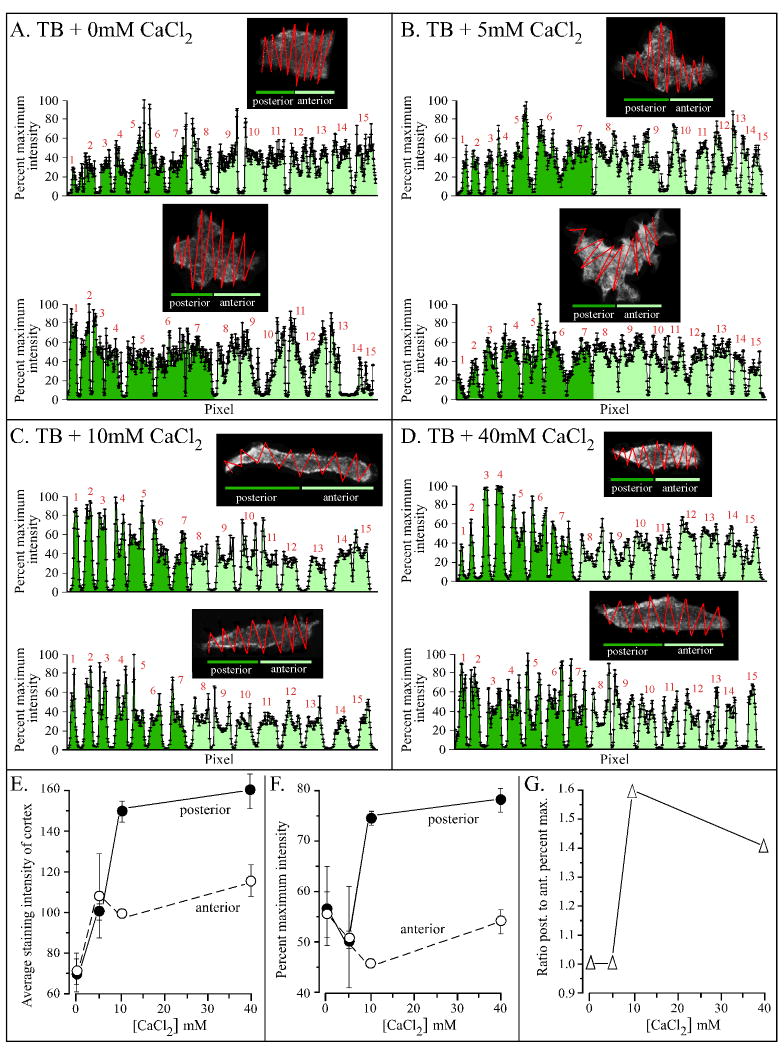
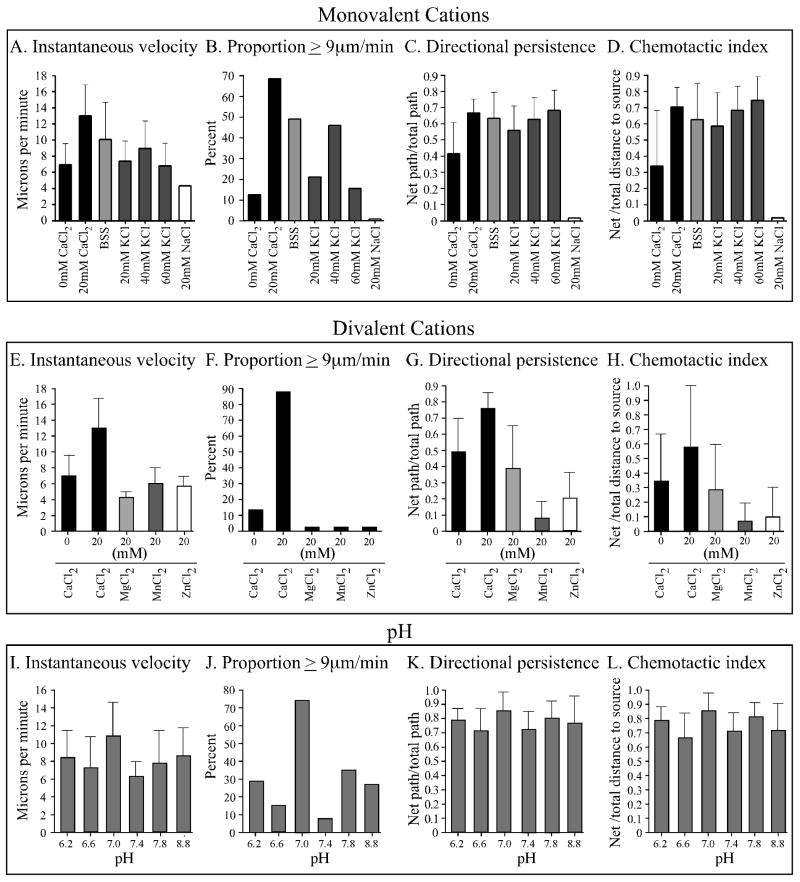
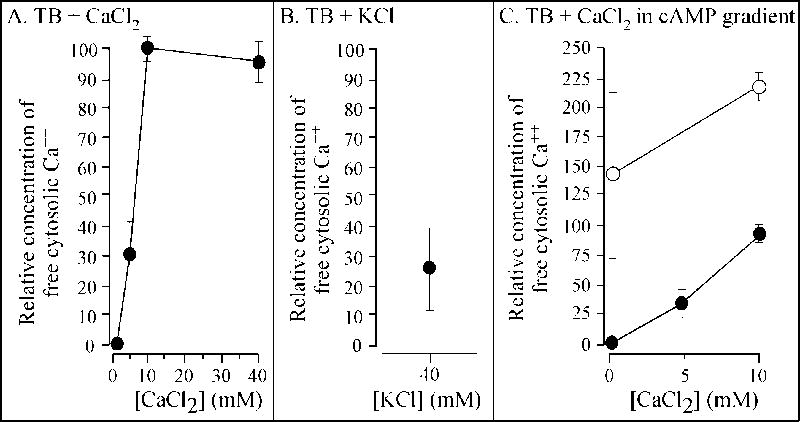
Extracellular Ca(++), a ubiquitous cation in the soluble environment of cells both free living and within the human body, regulates most aspects of amoeboid cell motility, including shape, uropod formation, pseudopod formation, velocity, and turning in Dictyostelium discoideum. Hence it affects the efficiency of both basic motile behavior and chemotaxis. Extracellular Ca(++) is optimal at 10 mM. A gradient of the chemoattractant cAMP generated in the absence of added Ca(++) only affects turning, but in combination with extracellular Ca(++), enhances the effects of extracellular Ca(++). Potassium, at 40 mM, can partially substitute for Ca(++). Mg(++), Mn(++), Zn(++), and Na(+) cannot. Extracellular Ca(++), or K(+), also induce the cortical localization of myosin II in a polar fashion. The effects of Ca(++), K(+) or a cAMP gradient do not appear to be similarly mediated by an increase in the general pool of free cytosolic Ca(++). These results suggest a model, in which each agent functioning through different signaling systems, converge to affect the cortical localization of myosin II, which in turn effects the behavioral changes leading to efficient cell motility and chemotaxis. Cell Motil. Cytoskeleton 2009. (c) 2009 Wiley-Liss, Inc.
Figures
Similar articles
-
Sphingosine-1-phosphate plays a role in the suppression of lateral pseudopod formation during Dictyostelium discoideum cell migration and chemotaxis.Cell Motil Cytoskeleton. 2004 Dec;59(4):227-41. doi: 10.1002/cm.20035. Cell Motil Cytoskeleton. 2004. PMID: 15476260
-
PTEN plays a role in the suppression of lateral pseudopod formation during Dictyostelium motility and chemotaxis.J Cell Sci. 2007 Aug 1;120(Pt 15):2517-31. doi: 10.1242/jcs.010876. Epub 2007 Jul 10. J Cell Sci. 2007. PMID: 17623773
-
Diverse chemotactic responses of Dictyostelium discoideum amoebae in the developing (temporal) and stationary (spatial) concentration gradients of folic acid, cAMP, Ca(2+) and Mg(2+).Cell Motil Cytoskeleton. 2002 Sep;53(1):1-25. doi: 10.1002/cm.10052. Cell Motil Cytoskeleton. 2002. PMID: 12211112
-
Signal transduction and motility of Dictyostelium.Biosci Rep. 1995 Dec;15(6):445-62. doi: 10.1007/BF01204348. Biosci Rep. 1995. PMID: 9156575 Review.
-
Chemotaxis: navigating by multiple signaling pathways.Sci STKE. 2007 Jul 24;2007(396):pe40. doi: 10.1126/stke.3962007pe40. Sci STKE. 2007. PMID: 17652307 Review.
Cited by
-
Systematic evaluation of buffer influences on the development of Dictyostelium discoideum.Dev Genes Evol. 2016 Jan;226(1):27-35. doi: 10.1007/s00427-016-0528-5. Epub 2016 Jan 20. Dev Genes Evol. 2016. PMID: 26791868
-
Calreticulin and cancer.Pathol Oncol Res. 2013 Apr;19(2):149-54. doi: 10.1007/s12253-012-9600-2. Epub 2013 Feb 8. Pathol Oncol Res. 2013. PMID: 23392843 Review.
-
The Roles of Signaling in Cytoskeletal Changes, Random Movement, Direction-Sensing and Polarization of Eukaryotic Cells.Cells. 2020 Jun 10;9(6):1437. doi: 10.3390/cells9061437. Cells. 2020. PMID: 32531876 Free PMC article. Review.
-
Ion Signaling in Cell Motility and Development in Dictyostelium discoideum.Biomolecules. 2024 Jul 10;14(7):830. doi: 10.3390/biom14070830. Biomolecules. 2024. PMID: 39062545 Free PMC article. Review.
-
Rectified directional sensing in long-range cell migration.Nat Commun. 2014 Nov 6;5:5367. doi: 10.1038/ncomms6367. Nat Commun. 2014. PMID: 25373620 Free PMC article.
References
-
- Abe T, Maeda Y, Iijima T. Transient increase of the intracellular Ca++ concentration during chemotactic signal transduction in Dictyostelium discoideum cells. Differentiation. 1988;39(2):90–6. - PubMed
-
- Alcantara F, Monk M. Signal propagation during aggregation in the slime mould Dictyostelium discoideum. J Gen Microbiol. 1974;85(2):321–34. - PubMed
-
- Andrew N, Insall RH. Chemotaxis in shallow gradients is mediated independently of PtdIns 3-kinase by biased choices between random protrusions. Nat Cell Biol. 2007;9(2):193–200. - PubMed
-
- Bagorda A, Mihaylov VA, Parent CA. Chemotaxis: moving forward and holding on to the past. Thromb Haemost. 2006;95(1):12–21. - PubMed
-
- Bangerth F. Calcium-Related Physiological Disorders of Plants. Annual Review of Phytopathology. 1979;17(1):97–122.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
