

Clustering of syntaxin-1A in model membranes is modulated by phosphatidylinositol 4,5-bisphosphate and cholesterol
- PMID: 19364135
- PMCID: PMC2724070
- DOI: 10.1021/bi9003217
Clustering of syntaxin-1A in model membranes is modulated by phosphatidylinositol 4,5-bisphosphate and cholesterol
Abstract
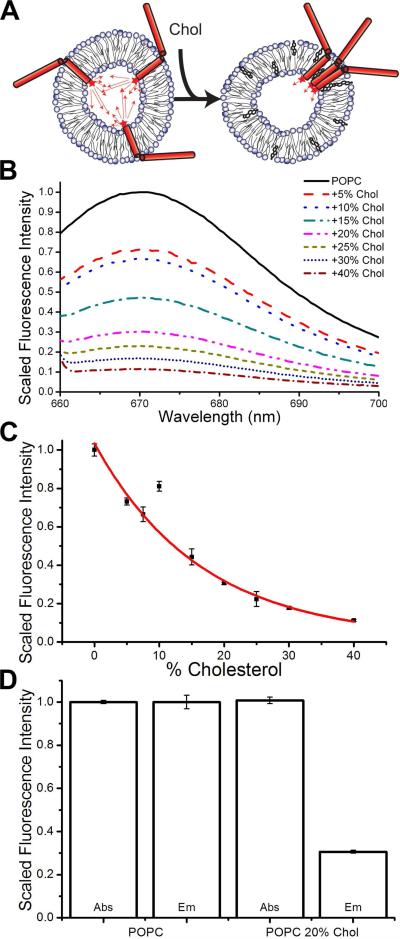
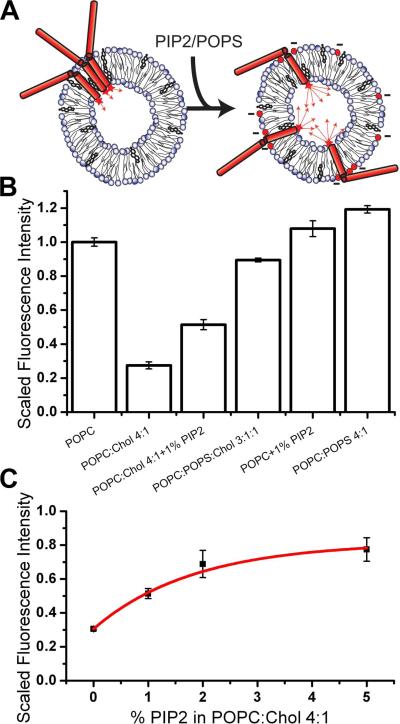
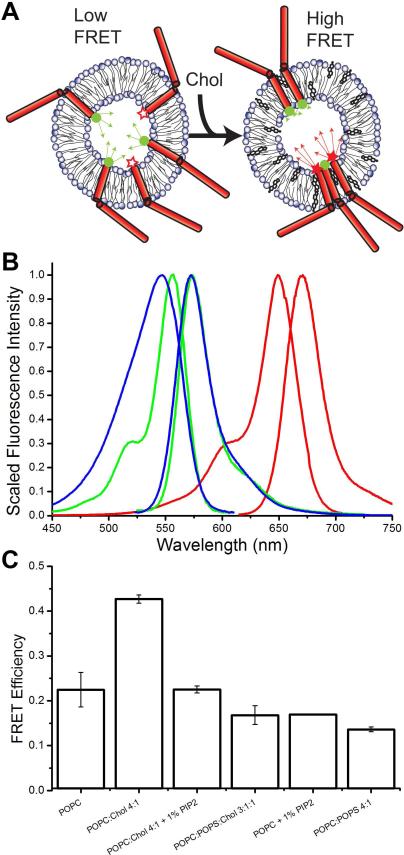
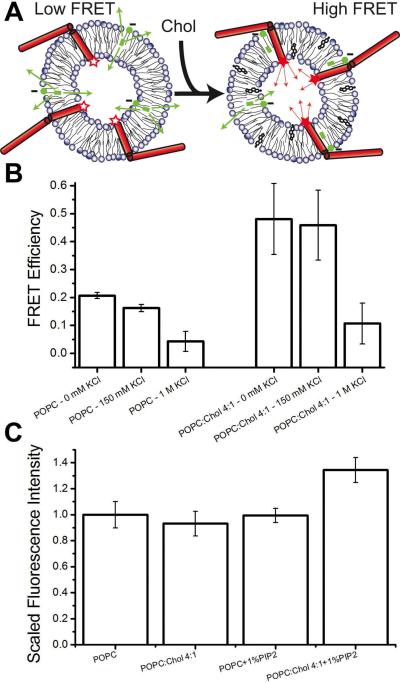
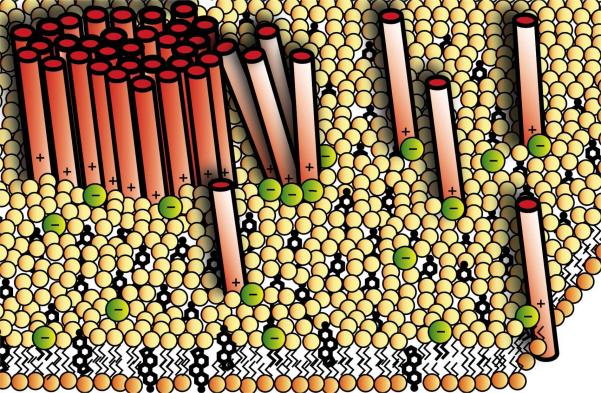
Syntaxin-1A is part of the SNARE complex that forms in membrane fusion in neuronal exocytosis of synaptic vesicles. Together with SNAP-25 the single-span transmembrane protein syntaxin-1A forms the receptor complex on the plasma membrane of neuroendocrine cells. Previous studies have shown that syntaxin-1A occurs in clusters that are different from lipid rafts in neuroendocrine plasma membranes. However, the interactions that promote these clusters have been largely unexplored. Here, we have reconstituted syntaxin-1A into lipid model membranes, and we show that syntaxin cluster formation depends on cholesterol in a lipid system that lacks sphingomyelin and therefore does not form liquid-ordered phases that are commonly believed to represent lipid rafts in cell membranes. Rather, the cholesterol-induced clustering of syntaxin is found to be reversed by as little as 1-5 mol % of the regulatory lipid phosphatidylinositol 4,5-bisphosphate (PI-4,5-P(2)), and PI-4,5-P(2) is shown to bind electrostatically to syntaxin, presumably mediated by the highly positively charged juxtamembrane domain of syntaxin. Possible implications of these results to the regulation of SNARE-mediated membrane fusion are discussed.
Figures
Similar articles
-
Membrane protein sequestering by ionic protein-lipid interactions.Nature. 2011 Oct 23;479(7374):552-5. doi: 10.1038/nature10545. Nature. 2011. PMID: 22020284 Free PMC article.
-
Molecular mechanism of cholesterol- and polyphosphoinositide-mediated syntaxin clustering.Biochemistry. 2011 Oct 25;50(42):9014-22. doi: 10.1021/bi201307u. Epub 2011 Sep 27. Biochemistry. 2011. PMID: 21916482 Free PMC article.
-
t-SNARE Transmembrane Domain Clustering Modulates Lipid Organization and Membrane Curvature.J Am Chem Soc. 2017 Dec 27;139(51):18440-18443. doi: 10.1021/jacs.7b10677. Epub 2017 Dec 13. J Am Chem Soc. 2017. PMID: 29231734 Free PMC article.
-
PI(4,5)P₂-binding effector proteins for vesicle exocytosis.Biochim Biophys Acta. 2015 Jun;1851(6):785-93. doi: 10.1016/j.bbalip.2014.09.017. Epub 2014 Oct 2. Biochim Biophys Acta. 2015. PMID: 25280637 Free PMC article. Review.
-
Need for speed: Super-resolving the dynamic nanoclustering of syntaxin-1 at exocytic fusion sites.Neuropharmacology. 2020 Jun 1;169:107554. doi: 10.1016/j.neuropharm.2019.02.036. Epub 2019 Feb 28. Neuropharmacology. 2020. PMID: 30826343 Review.
Cited by
-
Dynamical Organization of Syntaxin-1A at the Presynaptic Active Zone.PLoS Comput Biol. 2015 Sep 14;11(9):e1004407. doi: 10.1371/journal.pcbi.1004407. eCollection 2015 Sep. PLoS Comput Biol. 2015. PMID: 26367029 Free PMC article.
-
Supported double membranes.J Struct Biol. 2009 Oct;168(1):183-9. doi: 10.1016/j.jsb.2009.02.008. Epub 2009 Feb 21. J Struct Biol. 2009. PMID: 19236921 Free PMC article.
-
Reconstituting SNARE-mediated membrane fusion at the single liposome level.Methods Cell Biol. 2015;128:339-63. doi: 10.1016/bs.mcb.2015.02.005. Epub 2015 Apr 8. Methods Cell Biol. 2015. PMID: 25997356 Free PMC article.
-
The cortical acto-Myosin network: from diffusion barrier to functional gateway in the transport of neurosecretory vesicles to the plasma membrane.Front Endocrinol (Lausanne). 2013 Oct 21;4:153. doi: 10.3389/fendo.2013.00153. Front Endocrinol (Lausanne). 2013. PMID: 24155741 Free PMC article. Review.
-
Membrane protein sequestering by ionic protein-lipid interactions.Nature. 2011 Oct 23;479(7374):552-5. doi: 10.1038/nature10545. Nature. 2011. PMID: 22020284 Free PMC article.
References
-
- Jahn R, Scheller RH. SNAREs--engines for membrane fusion. Nat Rev Mol Cell Biol. 2006;7:631–643. - PubMed
-
- Weber T, Zemelman BV, McNew JA, Westermann B, Gmachl M, Parlati F, Sollner TH, Rothman JE. SNAREpins: minimal machinery for membrane fusion. Cell. 1998;92:759–772. - PubMed
-
- Bai J, Chapman ER. The C2 domains of synaptotagmin--partners in exocytosis. Trends Biochem Sci. 2004;29:143–151. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
