

Mechanisms controlling anaemia in Trypanosoma congolense infected mice
- PMID: 19365556
- PMCID: PMC2664899
- DOI: 10.1371/journal.pone.0005170
Mechanisms controlling anaemia in Trypanosoma congolense infected mice
Abstract
Background: Trypanosoma congolense are extracellular protozoan parasites of the blood stream of artiodactyls and are one of the main constraints on cattle production in Africa. In cattle, anaemia is the key feature of disease and persists after parasitaemia has declined to low or undetectable levels, but treatment to clear the parasites usually resolves the anaemia.
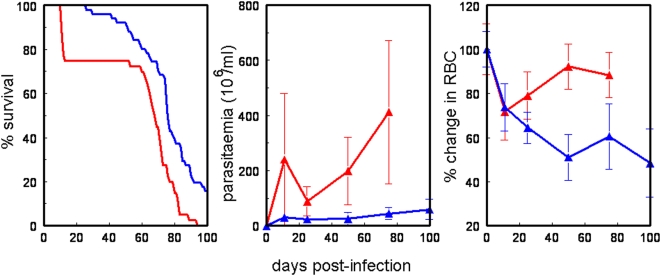
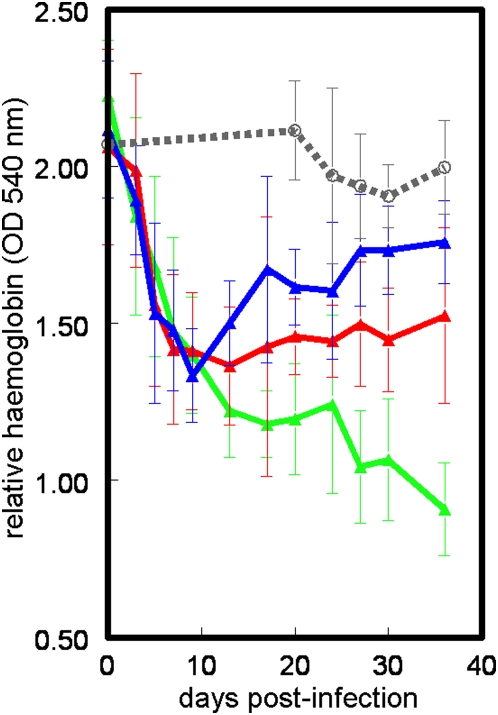
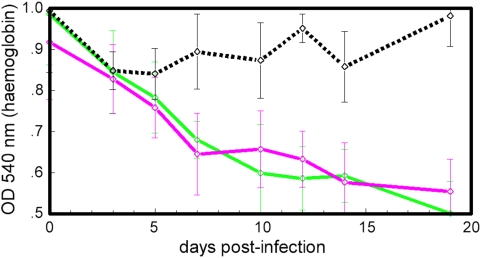
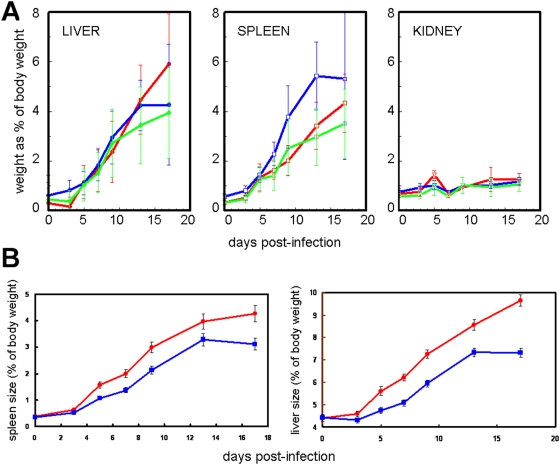
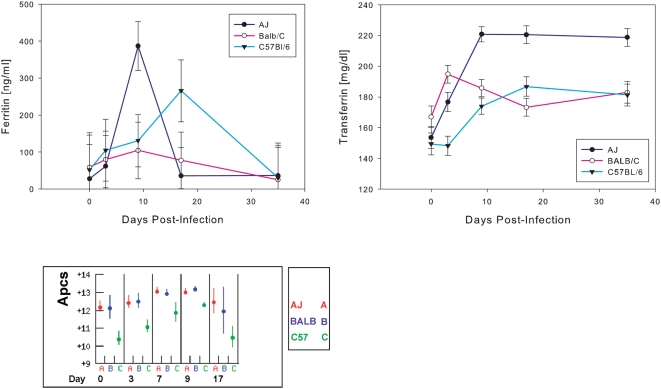
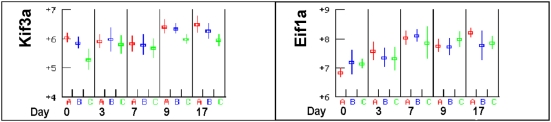
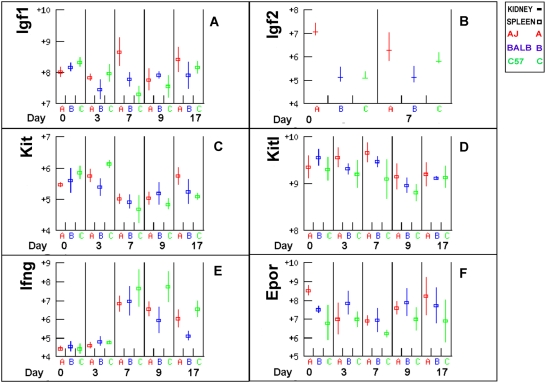
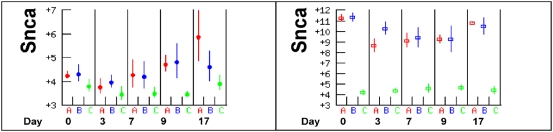
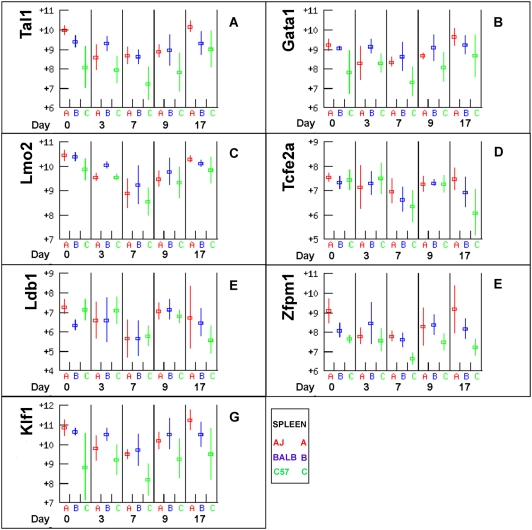
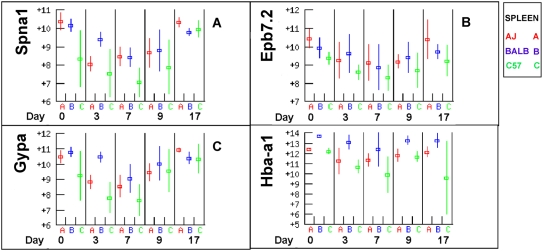
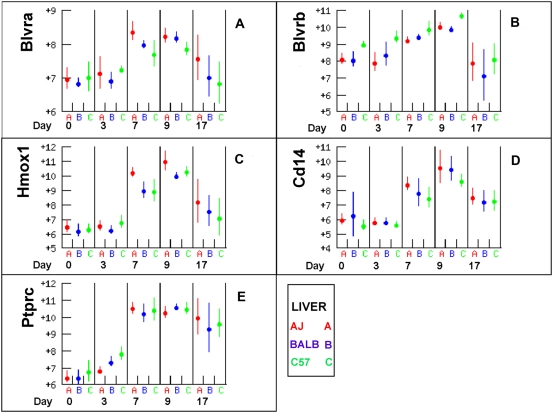
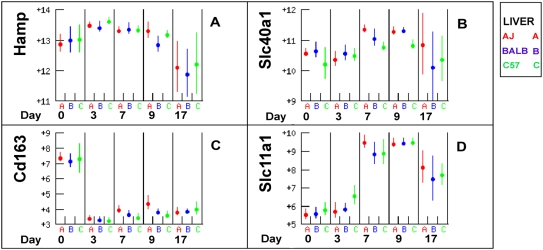
Methodology/principal findings: The progress of anaemia after Trypanosoma congolense infection was followed in three mouse strains. Anaemia developed rapidly in all three strains until the peak of the first wave of parasitaemia. This was followed by a second phase, characterized by slower progress to severe anaemia in C57BL/6, by slow recovery in surviving A/J and a rapid recovery in BALB/c. There was no association between parasitaemia and severity of anaemia. Furthermore, functional T lymphocytes are not required for the induction of anaemia, since suppression of T cell activity with Cyclosporin A had neither an effect on the course of infection nor on anaemia. Expression of genes involved in erythropoiesis and iron metabolism was followed in spleen, liver and kidney tissues in the three strains of mice using microarrays. There was no evidence for a response to erythropoietin, consistent with anaemia of chronic disease, which is erythropoietin insensitive. However, the expression of transcription factors and genes involved in erythropoiesis and haemolysis did correlate with the expression of the inflammatory cytokines Il6 and Ifng.
Conclusions/significance: The innate immune response appears to be the major contributor to the inflammation associated with anaemia since suppression of T cells with CsA had no observable effect. Several transcription factors regulating haematopoiesis, Tal1, Gata1, Zfpm1 and Klf1 were expressed at consistently lower levels in C57BL/6 mice suggesting that these mice have a lower haematopoietic capacity and therefore less ability to recover from haemolysis induced anaemia after infection.
Conflict of interest statement
Figures
Similar articles
-
Lack of correlation of anaemia with splenomegaly and hepatomegaly in Trypanosoma brucei and Trypanosoma congolense infections of rats.J Comp Pathol. 1997 Oct;117(3):261-5. doi: 10.1016/s0021-9975(97)80020-5. J Comp Pathol. 1997. PMID: 9447486
-
TNF-alpha mediates the development of anaemia in a murine Trypanosoma brucei rhodesiense infection, but not the anaemia associated with a murine Trypanosoma congolense infection.Clin Exp Immunol. 2005 Mar;139(3):405-10. doi: 10.1111/j.1365-2249.2004.02717.x. Clin Exp Immunol. 2005. PMID: 15730385 Free PMC article.
-
Bovine trypanotolerance: A natural ability to prevent severe anaemia and haemophagocytic syndrome?Int J Parasitol. 2006 May 1;36(5):521-8. doi: 10.1016/j.ijpara.2006.02.012. Epub 2006 Mar 13. Int J Parasitol. 2006. PMID: 16678182 Review.
-
Diminazene aceturate (Berenil) modulates the host cellular and inflammatory responses to Trypanosoma congolense infection.PLoS One. 2012;7(11):e48696. doi: 10.1371/journal.pone.0048696. Epub 2012 Nov 7. PLoS One. 2012. PMID: 23144931 Free PMC article.
-
Alternative versus classical macrophage activation during experimental African trypanosomosis.Int J Parasitol. 2001 May 1;31(5-6):575-87. doi: 10.1016/s0020-7519(01)00170-9. Int J Parasitol. 2001. PMID: 11334945 Review.
Cited by
-
The pathogenicity of blood stream and central nervous system forms of Trypanosoma brucei rhodesiense trypanosomes in laboratory mice: a comparative study.F1000Res. 2023 Nov 15;11:260. doi: 10.12688/f1000research.75518.2. eCollection 2022. F1000Res. 2023. PMID: 38162635 Free PMC article.
-
Genotype and expression analysis of two inbred mouse strains and two derived congenic strains suggest that most gene expression is trans regulated and sensitive to genetic background.BMC Genomics. 2010 Jun 7;11:361. doi: 10.1186/1471-2164-11-361. BMC Genomics. 2010. PMID: 20529291 Free PMC article.
-
Trypanosoma carassii infection in goldfish (Carassius auratus L.): changes in the expression of erythropoiesis and anemia regulatory genes.Parasitol Res. 2019 Apr;118(4):1147-1158. doi: 10.1007/s00436-019-06246-5. Epub 2019 Feb 12. Parasitol Res. 2019. PMID: 30747294
-
A polymorphism in the haptoglobin, haptoglobin related protein locus is associated with risk of human sleeping sickness within Cameroonian populations.PLoS Negl Trop Dis. 2017 Oct 27;11(10):e0005979. doi: 10.1371/journal.pntd.0005979. eCollection 2017 Oct. PLoS Negl Trop Dis. 2017. PMID: 29077717 Free PMC article.
-
Iron Homeostasis and Trypanosoma brucei Associated Immunopathogenicity Development: A Battle/Quest for Iron.Biomed Res Int. 2015;2015:819389. doi: 10.1155/2015/819389. Epub 2015 May 18. Biomed Res Int. 2015. PMID: 26090446 Free PMC article. Review.
References
-
- Kristjanson PM, Swallow BM, Rowlands GJ, Kruska RL, de Leeuw PN. Measuring the costs of African animal trypanosomosis, the potential benefits of control and returns to research. Agricultural Systems. 1999;59:79–98.
-
- Trail JCM, Dieteren GDM, Viviani P, Yangari G, Nantulya VM. Relationships between trypanosome infection measured by antigen-detection enzyme immunoassays, anemia and growth in trypanotolerant Ndama cattle. Veterinary Parasitology. 1992;42:213–223. - PubMed
-
- Naessens J, Leak SGA, Kennedy DJ, Kemp SJ, Teale AJ. Responses of bovine chimaeras combining trypanosomosis resistant and susceptible genotypes to experimental infection with Trypanosoma congolense. Veterinary Parasitology. 2003;111:125–142. - PubMed
-
- Naessens J. Bovine trypanotolerance: A natural ability to prevent severe anaemia and haemophagocytic syndrome? International Journal for Parasitology. 2006;36:521–528. - PubMed
-
- Murray M, Morrison WI, Whitelaw DD. Host susceptibility to African trypanosomiasis: trypanotolerance. Adv Parasitol. 1982;21:1–68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
