

Disulfide bond formation at the C termini of vaccinia virus A26 and A27 proteins does not require viral redox enzymes and suppresses glycosaminoglycan-mediated cell fusion
- PMID: 19369327
- PMCID: PMC2698572
- DOI: 10.1128/JVI.02295-08
Disulfide bond formation at the C termini of vaccinia virus A26 and A27 proteins does not require viral redox enzymes and suppresses glycosaminoglycan-mediated cell fusion
Abstract
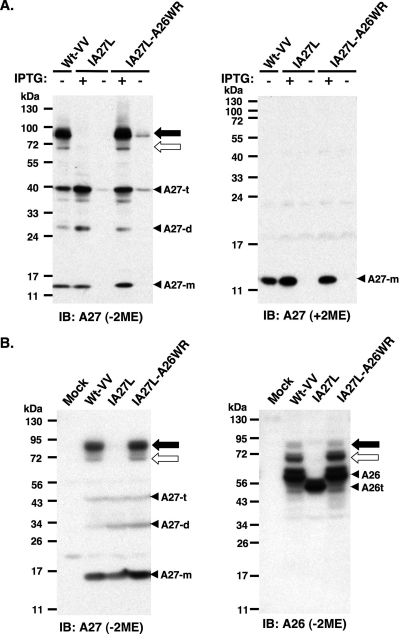
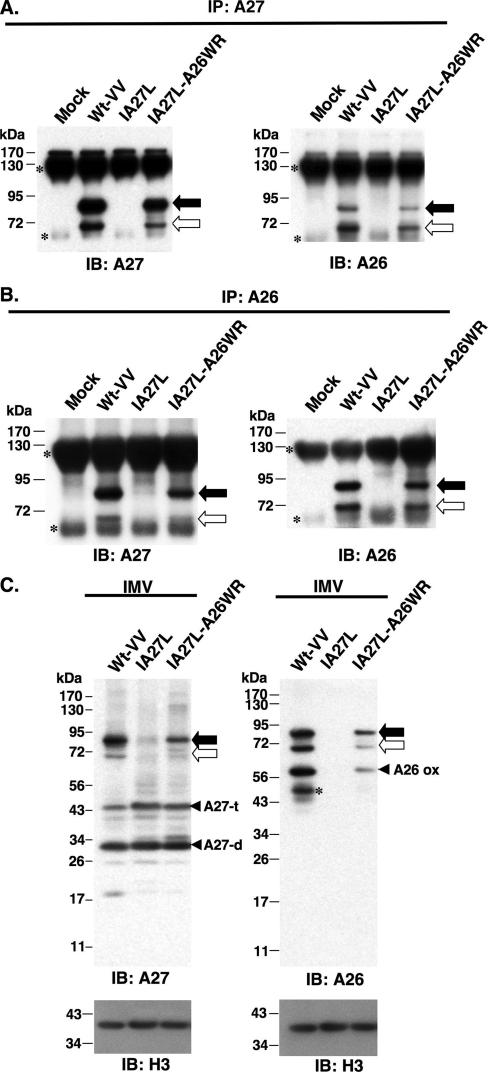
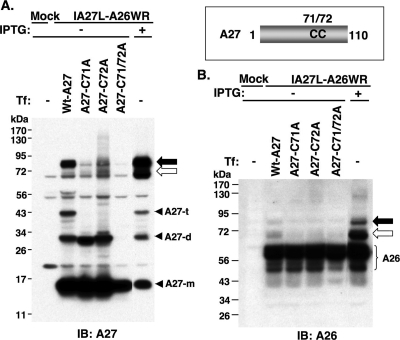
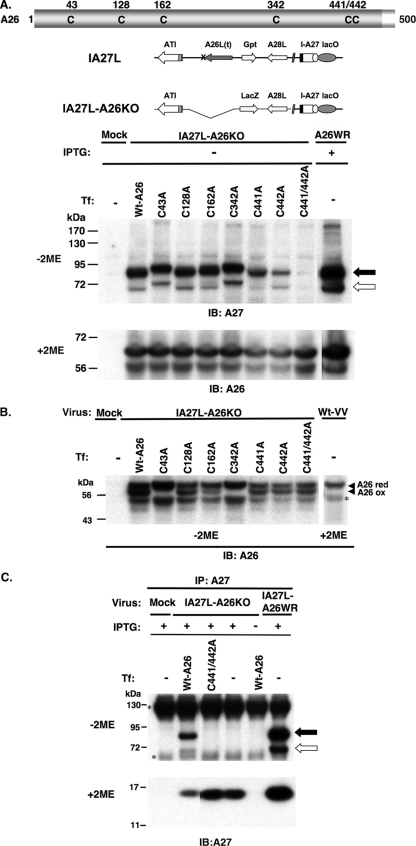
Vaccinia virus A26 protein is an envelope protein of the intracellular mature virus (IMV) of vaccinia virus. A mutant A26 protein with a truncation of the 74 C-terminal amino acids was expressed in infected cells but failed to be incorporated into IMV (W. L. Chiu, C. L. Lin, M. H. Yang, D. L. Tzou, and W. Chang, J. Virol 81:2149-2157, 2007). Here, we demonstrate that A27 protein formed a protein complex with the full-length form but not with the truncated form of A26 protein in infected cells as well as in IMV. The formation of the A26-A27 protein complex occurred prior to virion assembly and did not require another A27-binding protein, A17 protein, in the infected cells. A26 protein contains six cysteine residues, and in vitro mutagenesis showed that Cys441 and Cys442 mediated intermolecular disulfide bonds with Cys71 and Cys72 of viral A27 protein, whereas Cys43 and Cys342 mediated intramolecular disulfide bonds. A26 and A27 proteins formed disulfide-linked complexes in transfected 293T cells, showing that the intermolecular disulfide bond formation did not depend on viral redox pathways. Finally, using cell fusion from within and fusion from without, we demonstrate that cell surface glycosaminoglycan is important for virus-cell fusion and that A26 protein, by forming complexes with A27 protein, partially suppresses fusion.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
