

Role of the lymphoreticular system in prion neuroinvasion from the oral and nasal mucosa
- PMID: 19369351
- PMCID: PMC2698571
- DOI: 10.1128/JVI.00018-09
Role of the lymphoreticular system in prion neuroinvasion from the oral and nasal mucosa
Abstract
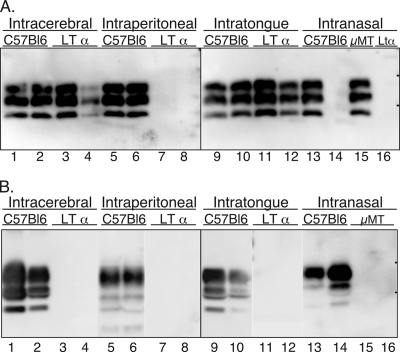
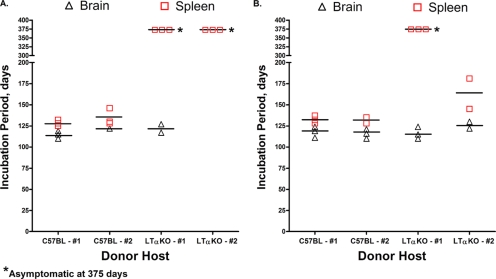
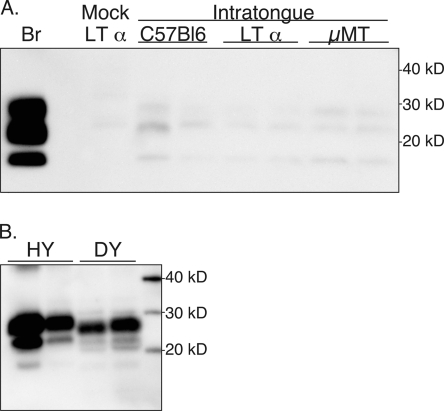
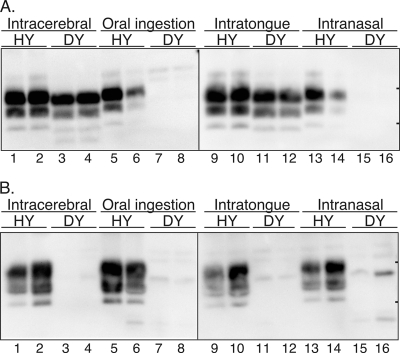
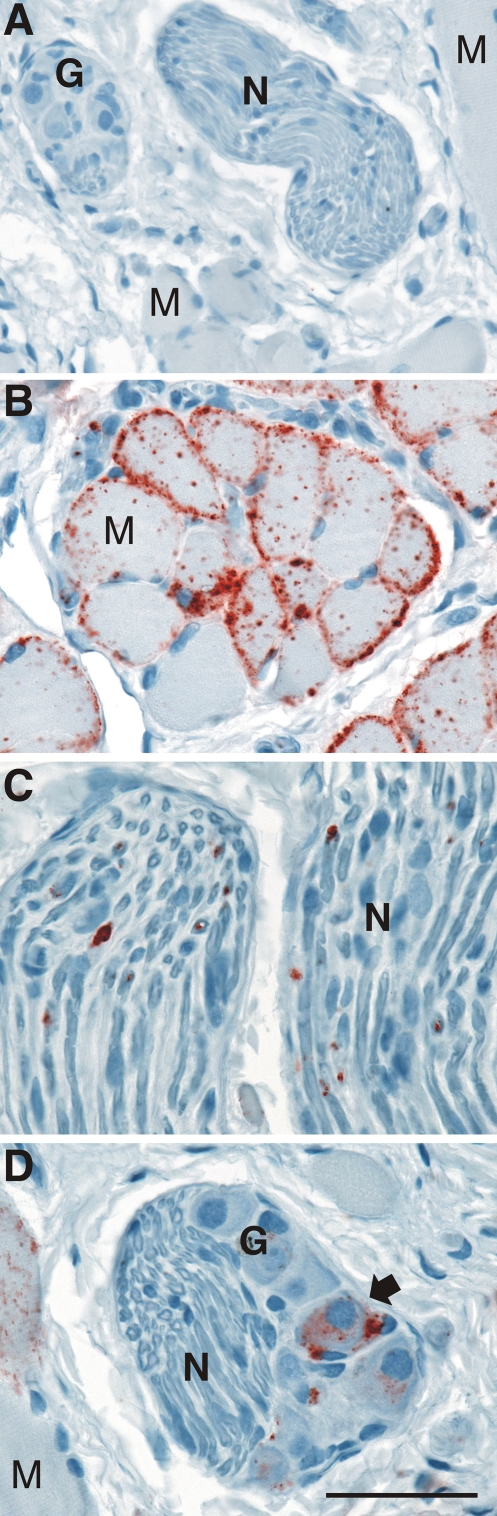
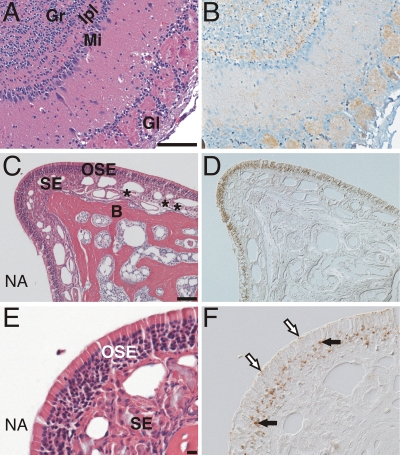
Prion neuroinvasion from peripheral tissues involves agent replication in the lymphoreticular system (LRS) prior to entry into the nervous system. This study investigated the role of the LRS in prion neuroinvasion from the oral and nasal mucosa in wild-type and immunodeficient mice and in hamsters infected with the HY and DY strains of the transmissible mink encephalopathy (TME) agent. Following inoculation at neural sites, all hosts were susceptible to prion disease and had evidence of prion infection in the brain, but infection of the LRS was found only in scrapie-infected wild-type mice and HY TME-infected hamsters. In the LRS replication-deficient models, prion neuroinvasion was not observed following intraperitoneal or oral inoculation. However, immunodeficient mice, which have impaired follicular dendritic cells, were susceptible to scrapie following intratongue and intranasal inoculation despite the absence of PrP(Sc) in the tongue or the nasal cavity. For DY TME, hamsters were susceptible following intratongue but not intranasal inoculation and PrP(Sc) was limited to nerve fibers of the tongue. These findings indicate that neuroinvasion from the tongue and nasal cavity can be independent of LRS infection but neuroinvasion was partially dependent on the strain of the prion agent and/or the host species. The paucity of PrP(Sc) deposition in the oral and nasal mucosa from LRS replication-deficient hosts following neuroinvasion from these tissues suggests an infection of nerve fibers that is below the threshold of PrP(Sc) detection and/or the transport of the prion agent along cranial nerves without agent replication.
Figures
References
-
- Banks, T. A., B. T. Rouse, M. K. Kerley, P. J. Blair, V. L. Godfrey, N. A. Kuklin, D. M. Bouley, J. Thomas, S. Kanangat, and M. L. Mucenski. 1995. Lymphotoxin-alpha-deficient mice. Effects on secondary lymphoid organ development and humoral immune responsiveness. J. Immunol. 1551685-1693. - PubMed
-
- Bartz, J. C., J. M. Aiken, and R. A. Bessen. 2004. Delay in onset of prion disease for the HY strain of transmissible mink encephalopathy as a result of prior peripheral inoculation with the replication-deficient DY strain. J. Gen. Virol. 85265-273. - PubMed
-
- Benestad, S. L., P. Sarradin, B. Thu, J. Schonheit, M. A. Tranulis, and B. Bratberg. 2003. Cases of scrapie with unusual features in Norway and designation of a new type, Nor98. Vet. Rec. 153202-208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
